

История развития учения о биологическом окислении
Еще древние философы отмечали взаимосвязь между процессами жизнедеятельности и дыханием. Они также провели параллель между дыханием и горением.
Платон утверждал, что воздух нужен для охлаждения внутреннего жара сгорающего вещества.
Аристотель утверждал, что воздух нужен для поддержания внутреннего горения.
В XVII – XVIII вв широкое признание получила теория горючего начала – флогистона, сформулированная Штаммом. Эта теория объясняла процессы горения выделениями их горящего тела особого невесомого в-ва, и была опровергнута Ломоносовым и Лавуазье, которые открыли закон сохранения энергии.
В XVIII веке с развитием физики газов, с появлением соответствующей аппарутуры стали проводить опыты по сжиганию веществ в замкнутом пространстве. В это время Шталем была сформулирована теория флагистона (горючего начала), согласно которой все вещества, подвергающиеся окислению состоят из оксида и флагистона.
В середине XVIII века было установлено:
1) процесс горения идет в воздушной среде с высокой температурой, дыхание – в среде с пониженной температурой;
2) при дыхании, как и при горении выделяется тепло, но в незначительных количествах;
3) конечные продукты CO2 и H2O.
В 1751 году Ломоносов подробно рассмотрел процессы горения и окисления.
В 1774 году Лавуазье повторил опыты Ломоосова и показал, что процессы горения и дыхания идентичны, т. к. образуются идентичные продукты.
Лавуазье назвал дыхание медленным горением и показал процесс сгорания Гл в организме:
C6H12O6 6O2 ——> 6CO2 6H2O Q
В начале XIX века стали известными катализаторы, с помощью которых осуществлялись процессы окисления. Это были металлы, обладающие «внутренней силой».
В середине XIX века немецкий ученый Шейнбайн, открывший озон, предположил, что в организме образуется озон и он используется в реакциях окисления.
После работ Лавуазье в науке господствовало мнение о тождестве горения и медленного окисления питательных вкществ в организме. Вместе с тем было ясно, что БО протекает в необычных условиях:
– при пониженной температуре;
– без пламени;
– и в присутсвии большого количества H2O (75% – 80% ткани).
В XIX веке появилось понятие о ферментах и причину своеобразного течения БО попытались объяснить «активацией» кислорода в клетках организма.
Одна из теорий была выдвинута Бахом, который считал, что «активация» молекулярного кислорода происходит в результате разрыва связи и присоединения к ферментам оксигеназам (А), способным к аутооксидации:
O A O2 —–> A |
O перекись
O
A | SH2 —–> S A H2O2 O субстарат
3 положения Баха:
1. Наличие высокоактивной оксидазы, но это не было обнаружено.
2.В тканях должна быть высокая концентрация H2O2, но это тоже не было обнаружено.
3. Высокая активность ферментов, разлагающих H2O2; это было обнаружено, существует 2 фермента:
каталаза
2H2O2 ————> 2H2O O2
Существует и другой механизм разложения H2O2:
2GSH H2O2 —–> 2H2O (пероксидаза) или глутатион |
SH2 H2O2 —–> S 2H2O —
Эта теория да и все остальные основывались на неправильном представлении об ОВР. Окислительный процесс рассматривался как процесс взаимодействия любого вещества с кислородом. То есть кислород – это окислитеоь.
К концу XIX века с разхвитием физики ядра и накопления знаний о структуре вещества, было установлено, что не все процессы окисления требуют для своей реализации наличие кислорода.
Кроме этого теория Баха основывалась на том, что в организме имеется большое количество ароматических соединений, на самом же деле их очень мало, в оновном глюкоза.
Согласно современных представлений ОВР – это процесс перемещения электронов и протонов от донора (восстановителя) – это процесс окисления – к акцептору (окислителю) – процесс восстановления.
Количественной мерой ОВР является величина ОВП. В начале точки отсчета стандартного потенциала взят ОВП водорода.
В 1912 году была сформулирована теория Палладина-Виланда, согласно которой в организме есть промежуточные вещества, способные акцептировать электроны и протоны от субстрата с последующей передачей электронов и протонов на кислород, по этой теории весь процесс БО можно разбить на 2 этапа:
1) анаэробный – передача электронов и протонов с субстрата на промежуточное вещество;
2) аэробный – передача электронов и протонов с промежуточного вещества на кислород.
Палладин предпологал, что существует несколько промежуточных переносчиков, позволяющих организму поэтапно освобождать химическую энергию и кислород выступает в качестве конечного акцептора электронов и протонов.
1 анаэробный этап:
SH2 —-> S
2 аэробный этап:
½ O2 —–> H2O
Роль промежуточных переносчиков (хромогенов) выполняют коферменты (НАД; НАДФ; ФАД; ФМН) оксидоредуктаз.
В последствии развития учения о БО, шло по пути развития знаний о хромогенах.
В 1925 году были открыты гистогематины (цитохромы).
В 1932 году академик Энгельгардт показал, что процесс окисления идет с образованием АТФ (окислительное фосфорилирование).
В 1945 году Ленинджер и Кенеди впервые показали, что процесс окисления веществ, цикл Кребса локализованы в митохондриях.
Современные представления о БО базируются на сущности трактовки ОВП, а также БО основано на общих законах термодинамики:
1 закон – закон сохранения энергии: энергия никуда не исчезает, а только переходит из одной формы в другую, т. е. сохраняется.
2 закон – все тела и химические процессы стремятся к минимуму энергии, к состоянию покоя и беспорядка, т. е. к энтропии.
С термодинамической точки зрения – организм человека – антиэнтропийная машина, открытая система, которая обменивается с окружающей средой веществом и энергией. Основа ее жизнедеятельности – обмен веществ (метаболизм).
§
Обмен веществ и энергии – закономерный порядок превращения вещества и энергии в живых организмах, направленный на их сохранение и самовоспроизведение. Обмен веществ и обмен энергии тесно связаны и представляют собой диалектическое единство.
Вся совокупность химических реакций, протекающих в живых органихмах, включая усвояемость веществ, поступающих извне (ассимиляция) и их расщепление (диссимиляция) вплоть до образования конечных продуктов, подлежащих выделению, составляет сущность и содержание обмена веществ.
Ассимиляция – одна из сторон обмена веществ.
Ассимиляция включает огромное количество химических превращений, приводящих к использованию органических и неорганических веществ, поступающих из внешней среды для построения специфических для данного организма белков, НК, липидов, углеводов. Процесс ассимиляции обеспечивает рост, развитие, обновление организма и накопление запасов, используемых в качестве источника энергии.
Диссимиляция – противоположная ассимиляции сторона обмена веществ: разрушение органических соединений с превращением их в простые вещества (в основном H2O, CO2, NH3).
Промежуточный обмен – превращение веществ в организме с момента поступления их в клетки до образования конечных продуктов.
Попав внутрь клетки, питательное вещество метаболизируется – претерпевают ряд химических изменений, катализируемых ферментами. Определенная последовательность таких химических изменений называется метаболическим путем, а образующиеся прмежуточные продукты – метаболитами.
Различают 2 стороны промежуточного обмена – анаболизм и катоболизм. Анаболические реакции направлены на образование и обновление структурных элементов клеток и тканей. Эти реакции преимущественно восстановительные, соправаждаются затратой свободной энергии.
Катоболические превращения – процессы расщепления сложных молекул, как поступивших с пищей, так и входящих в состав клетки до простых компонентов. Эти реакции обычно окислительные, сопроваждаются выделением свободной энергии.
Обе стороны промежуточного обмена тесно взаимосвязаны во времени и пространстве.
Боилогическое окисление – это совокупность биохимических реакций, приводящих к образованию полезной энергии за счет деградации компонентов пищи.
Принципиальной особенностью БО или тканевого дыхания является то, что оно протекает постепенно, через многочисленные промежуточные стадии, т. е. происходит многократная передача протонов и электронов от донора к акцептору.
Субстраты биологического окисления.
Субстратом БО является любое вещество, способное поставлять электроны и протоны, энергия которых трансформируется в полезную конвертируемую форму.
Субстраты БО: метаболиты восстанавливающие НАД, ФАД, служащие предшественниками субстратов, зависящие от дегидрогеназ Гл, АК.
Схема энергетического обмена.
Основные компоненты пищи – белки, липиды и углеводы проходят 3 этапа энергетического обмена:
1. ЖКТ – происходит деполимеризация сложных соединений: крахмал и гликоген —–> Гл
дисахариды и олиго —–> до моносахаридов
белки —–> до пептидов и АК.
2. С момента поступления мономеров в клетку начинается цитозольный этап: происходит дальнейший распад мономеров и унификация субстратов, превращение их в общие соединения:
Углеводы все идут в Гл.
Липиды —-> Гн и ЖК.
3. Митохондриальный – унификация субстратов продолжается в митохондриальном матриксе, тут субстраты подвергаются окислению путем вовлечения в цикл Кребса, который снимает с них электроны и протоны и трансформирует их энергию в конвертируемую форму АТФ.
Схема образования субстратов биологического окисления.
БЕЛКИ УГЛЕВОДЫ ЛИПИДЫ Энергия
| | | | |
АК Гл Гн ЖК |1; 0,5%
| ЩУК | | 3ФГА | | ПВК — | | АцКоА | > | лактат | | | | |2; 2,5% | | | | | |
| Цикл Кребса NAD 1/2O2 ——– АДФ Фн | | Цитрат NAD.H2 -> H2O АТФ работа | —————– | | | | | | 3; 97% | | | ———- |
§
Роль АТФ – хранилище биологической энергии. В 1 молекуле АТФ имеется 2 макроэргические связи. При их расщеплении высвобождается 32 кДж энергии.
АТФ присутствует в клетках в диссоциированной форме: АТФ4-АТФ4- ——> АДФ3- Фн2- Н
10-3 10-3 10-3 10-7
Т. о. всякая работа в клетке сопровождается образованием H , которые захватываются буферами.
1 причина макроэргичности: т. к. концентрация АТФ, АДФ и Фн одинакова (по 10-3 моль), а концентрация Н = 10-7 моль,
согласно закону соотношения действующих масс равновесие сдвинуто вправо.
2 причина: в структуре АТФ имеется 3 фосфата и 2 ангидридные связи, за счет этого на хвосте молекулы АТФ создается конфармационная напряженность, возникает сила электростатического отталкивания и АТФ отдает молекулу фосфата. И при этом она переходит в более выгодное состояние АДФ Фн, которое более устойчиво, это 3-я причина макроэргичности.
В клетках АТФ присутствует в виде магниевой соли. Существует точка зрения, что уровень Mg2 отражает уровень АТФ.
В 1940-41 гг немецким биохимиком Фрицем Липманом была создана концепция АТФ-азного цикла: в процессе фото- или хемосинтеза энергия депонируется в форме АТФ. Синтез АТФ в организме происходит из АМФ:
Фн
АМФ Фн ——> АДФ ——-> АТФ
HOH
фото -> АТФ ———-> осмотическая работа
синтез | —-> транспорт
хемо —АДФ Фн<—–> электрическая работа
—-> химическая работа
—-> тепловая работа
—-> механическая и световая работа.
§
Такое «центральное» расположение молекулы АТФ позволяет ей выполнять роль донора высокоэнергетического фосфата для соединений, расположенных ниже в таблице, превращаясь при этом в АДФ, а АДФ – роль акцептора высокоэнергетического фосфата у соединений, расположенных выше. Цикл АТФ/АДФ связывает, тем самым, процессы генерирующие «~Р» с процессами, использующими «~Р». Сумму всех адениловых нуклеотидов в клетке (АТФ,АДФ и АМФ) называют адениловой системой. Процессы гидролиза и синтеза АТФ происходят с высокой скоростью, поскольку общий фонд АТФ очень маленький и для поддержания процессов жизнедеятельности в клетке его хватает только на несколько секунд.
В клетках организмов животных есть три основных источника ~P для синтеза АТФ.
· окислительное фосфорилирование – механизм образования АТФ, использующий для этого энергию градиента электрохимического потенциала, возникающего на внутренней мембране митохондрий.
· Субстратное фосфорилирование – механизм синтеза АТФ, использующий энергию макроэргических соединений, образующихся в процессе метаболизма (1,3- дифосфоглицериновая кислота, сукцинил-КоА и т.д.).
· Синтез АТФ с использованием макроэргов, выполняющих своеобразную роль молекул – депо макроэргических связей (креатинфосфат).
Тому, каким образом живые системы преобразуют энергию поступающих из внешней среды химических соединений, в энергию макроэргических соединений и посвящена значительная часть курса биохимии.
§
Цикл трикарбоновых кислот или цикл лимонной кислоты был открыт Гансом Кребсом в 1937 г. Он брал измельченные мышцы голубя, добавлял на них трикарбоновые кислоты и определял скорость дыхания, те трикарбоновые кислоты, которые составляют цикл Кребса усиливают дыхание.
Цикл Кребса – исходный субстрат ацетил КоА, который взаимодействует с ЩУК под действием фермента цитратсинтетазы.
За один оборот цикла Кребса происходит полное окисление одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление ацетил-КоА, а коферменты НАД и ФАД должны снова окисляться. Это и происходит в ДЦ.
Освобождающаяся при окислении ацетил-КоА энергия, расходуется на образование макроэргических связей АТФ.
Из 4 пар атомов водорода, 3 пары переносятся через НАД и одна пара через ФАД. На каждую пару атомов водорода в системе БО образуется 3АТФ (1НАДН2 = 1АТФ). Следовательно, всего 9АТФ; одна пара атомов попадает в систему БО через ФАД, – в результате образуется 2АТФ. Кроме этого в ходе сукцинаткиокиназной реакции образуется 1ГТФ = 1АТФ. Поэтому в целом, в ходе цикла Кребса образуется 12АТФ.
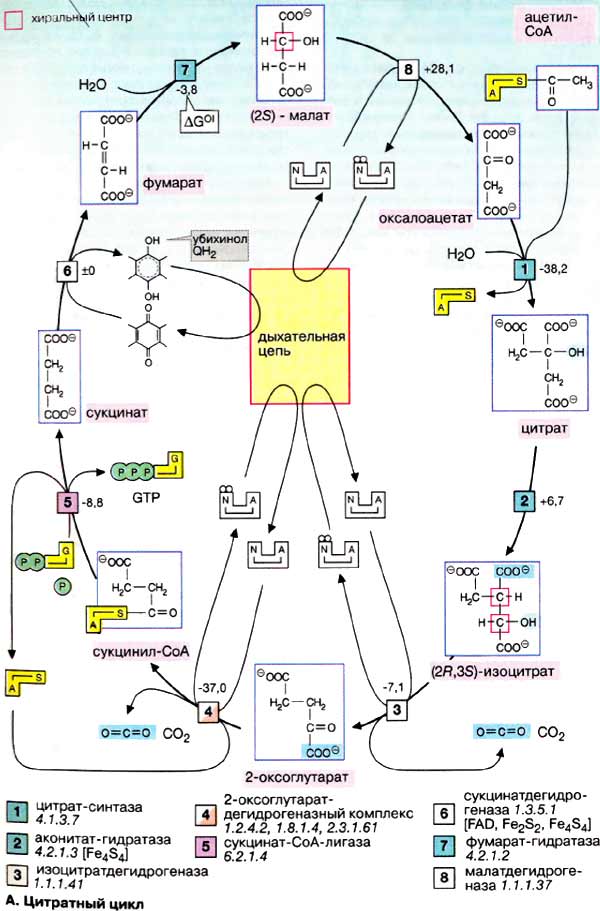
§
ЦТК связан с предшествующими стадиями энергетического обмена (гликолиз, окисление ЖК и АК), поэтому механизмы регуляции этих процессов будут справедливы и для ЦТК:
1) ретроингибирование; 2) путем изменения концентрации субстрата на входе ЦТК; 3) аллостерическаярегуляция (с помощью НАД, НАДН2, АТФ); 4) ионная (pH, [Ca ]).
Так как цикл Кребса начинается со стадии ЩУК ацетил КоА, то эти метаболиты и управляют интенсивностью ЦТК. Первым регуляторным фактором является концентрация ЩУК, которая в основном образуется из ПВК, ацетил КоА в принципе тоже:
CO2 -CO2
ЩУК <—– ПВК ——> ацетил КоА
ГТФ
ПВК же образуется из углеводов (Гл), поэтому при диабете или углеводном голодании наблюдается недостаток ПВК, а значит и ЩУК и ЦТК блокируются. Ацетил-КоА не является лимитирующим субстратом, т. к. в основном образуется при окислении ЖК.
Но в то же время ЩУК – конкурентный ингибитор сукцинатдегидрогеназы, поэтому при избытке ЩУК, ЦТК блокируется на 6 стадии (так называемое «щуковое торможение»). Это торможение можно убрать ГЛУ, который переаминирует ЩУК в АСП.
Второй регуляторный фактор – концентрация НАД и НАДН2. В живых системах концентрация НАД НАДН2 = const. Любые факторы, ведущие к увеличению НАД.Н2 (гипоксия, алкогольная интоксик5ация) и дефициту НАД блокирует ЦТК. Следовательно увеличение концентрации НАД при активной работе ДЦ стимулирует ЦТК.
Так как АТФ является косвенно конечным продуктом ЦТК, то ее избыток блокирует ЦТК, а значит АДФ стимулирует ЦТК. (АДФ рассматривается как аллостерический активатор изоцитратдегидрогеназы).
Стимулятором ЦТК является также кислород, потому что он стимулирует распад АТФ.
Нормальная концентрация Ca2 в клетке 10-7 моль. При увеличении концентрации кальция до 10-6 моль активируются дегидрогеназные реакции: пируватДГ, изоцитратДГ, альфа-КГДГ, а значит и ЦТК.
Цикл Кребса активируется при сердечной недостаточности. Это объясняется тем, что миокард не может самостоятельно убрать избыток Ca2 и эту роль берут на себя митохондрии, возрастает потребность в кислороде.
| Заведующий кафедрой биологической химии, д.м.н., проф. | Грицук А. И. | ___________ |
21.10.2006
Министерство здравоохранения Республики Беларусь
УО «Гомельский государственный медицинский университет»
Кафедра биологической химии
Обсуждено на заседании кафедры (МК или ЦУНМС)
Протокол № _________________200__года
ЛЕКЦИЯ
по биологической химии
наименование дисциплины
для студентов _2__ курса лечебного факультета
Тема Биологическое окисление 2. Тканевое дыхание. Окислительное фосфорилирование.
Время 90 мин.
Учебные и воспитательные цели:
Дать представление:
1. О строении дыхательной цепи (ДЦ), ингибиторах; механизмах работы ДЦ; пунктах сопряжения, величинах ОВП компонентов ДЦ. О коэффициенте Р/О, его значении.
2. О свободном и разобщенном дыхании. О теориях сопряжения ОФ.
3. О механизме генерации DmН .
4. О структуре и функциях протонной АТФ-азы; о механизме разобщения.
5. Об окислительдном фосфорилировании (DpH и Dy); о механизмах термогенеза, роли бурой жировой ткани.
6. О роли энергетического обмена; Путях утилизации DmН и АТФ. О прикладных аспектах биоэнергетики.
7. О путях потребления O2 в организме (митохондриальный, микросомальный, перекисный). О характеристике микросомальной ДЦ, в сравнении с митохондриальной. О характеристике цитохромаP450, функции.
8. О перекисном окисление. О механизме образования активных форм кислорода O2–, O2, O2 . О роли перекисных процессов в норме и при патологии. О перекисном окислении липидов (ПОЛ): (НЭЖК → R· → диеновые коньюгаты → гидроперекиси → МДА). О способах оценки активности ПОЛ.
9. Об антиоксидантной защите: ферментной и неферментной. О характеристиках СОД, каталазы, глютатионпероксидазы, GSH-редуктазы, NADPH-воспроизводящих систем. О неферментных АОС: витаминах Е, А, С, каротиноидах, гистидине, кортикостероидах, билирубине, мочевине и др.
ЛИТЕРАТУРА
1 Березов Т. Т., Коровкин Б. Ф. Биологическая химия. М.: Медицина, 1990. С. 213–220; 1998. С. 305–317.
2 Николаев А. Я. Биологическая химия. М.: Высшая школа, 1989. С. 199–221.
Дополнительная
3 Филиппович Ю. Б. Основы биохимии. М.: Высшая школа, 1993. С. 403–438.
4 Марри Р. и др. Биохимия человека. М.: Мир, 1993. Т. 1. С. 111–139.
5 Ленинджер А. Основы биохимии. М.: Мир, 1985. Т. 2. С. 403–438, 508–550.
6 АлбертсБ.идр.,Молекулярнаябиология клетки. М.: Мир, 1994.Т. 1. С. 430–459.
7 Скулачев В.П. Энергетика биологическихмембран. М.: Наука. 1989.
МАТЕРИАЛЬНОЕ ОБЕСПЕЧЕНИЕ
1. Мультимедийная презентация.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
| № п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
| 1. | Строение дыхательной цепи (ДЦ), ее комплексы, ингибиторы. Механизм работы ДЦ. Пункты сопряжения, величина ОВП компонентов ДЦ. Коэффициент Р/О, его значение. | |
| 2. | Свободное и разобщенное дыхание. Теории сопряжения ОФ (химическая, конформационная, хемиосмотическая – П. Митчелла). | |
| 3. | Механизм генерации DmН , его компоненты, стехиометрия Н /е. | |
| 4. | Структура и функция протонной АТФ-азы. Механизм разобщения. | |
| 5. | ОФ (снятие DpH и Dy). Механизмы термогенеза. Роль бурой жировой ткани. | |
| 6. | Основополагающая роль энергетического обмена. Пути утилизации DmН и АТФ. Прикладные аспекты биоэнергетики. | |
| 7. | Пути потребления O2 в организме (митохондриальный, микросомальный, перекисный). Характеристика микросомальной ДЦ, ее сравнение с митохондриальной. Характеристика цитохромов P450, их функция. | |
| 8. | Перекисное окисление. Механизм образования активных форм кислорода O2–, O2, O2 . Роль перекисных процессов в норме и при патологии. Общее представление о ПОЛ (НЭЖК → R· → диеновые коньюгаты → гидроперекиси → МДА). Способы оценки активности ПОЛ. | |
| 9. | Антиоксидантная защита: ферментная и неферментная. Характеристика СОД, каталазы, глютатионпероксидазы, GSH-редуктазы, NADPH-воспроизводящих систем. Неферментные АОС: витамины Е, А, С, каротиноиды, гистидин, кортикостероиды, билирубин, мочевина и др. |
Всего 90 мин
§
Поэтапное «контролируемое сгорание» достигается путём промежуточного включения дыхательных ферментов, обладающих различным редокс-потенциалом. Редокс-потенциал (окислительно-восстановительный потенциал) определяет направление переноса протонов и электронов ферментами дыхательной цепи (рис.1).
Редокс-потенциал выражается значением электродвижущей силы (в вольтах), которая возникает в растворе между окислителем и восстановителем, присутствующих в концентрации 1,0 моль/л при 25˚ С (при рН=7,0 оба находятся в равновесии с электродом, который может обратимо принимать электроны от восстановителя). При рН=7,0 редокс-потенциал системы Н2 /2Н 2ē равен – 0,42 v. Знак – означает, что данная редокс-пара легко отдаёт электроны, т.е. играет роль восстановителя, знак указывает на способность редокс-пары принимать электроны, т.е. играть роль окислителя. Например, редокс-потенциал пары НАДН∙Н / НАД равен – 0,32 v, что говорит о высокой её способности отдавать электроны, а окислительно-восстановительная пара ½О2 /Н2О имеет наибольшую положительную величину 0,81 v, т.е. кислород обладает наивысшей способностью принимать электроны.
В процессе окисления АцКоА в ЦТК, восстановленные формы НАДН2 и ФАДН2 поступают в ДЦ, где энергия электронов и протонов трансформируется в энергию макроэргических связей АТФ.
ДЦ – совокупность дегидрогеназ, которые транспортируют электороны и протоны с субстрата на кислород.
Принципы функционирования ДЦ основаны на 1-ом и 2-ом законах термодинамики.
Движущей силой ДЦ является разность ОВП. Суммарная разность всей ДЦ составляет 1,1 В. Пункты фосфорилирования должны иметь перепад ОВП = 0,25 – 0,3 В.
1. Пара НАД-Н имеет ОВП = 0,32 В.
2. Пара Q-b – / – /- – 0 В.
3. O2 – имеет 0,82 В.
ДЦ локализуется во внутренней мембране митохондрий и имеет 2 пути введения электронов и протонов или 2 входа; ДЦ образует 4 комплекса.
1 вход: НАД-зависимый (поступают электроны и протоны со всех НАД-зависимых реакций).
2 вход: ФАД-зависимый
НАД —->ФП
Q —>b—>c1—>c—>aa3—->1/2O2
Янтарная кислота —->ФП
Дыхательная цепь – форма реализации биологического окисления.
Тканевое дыхание – это последовательность окислительно-восстанови-тельных реакций, протекающих во внутренней митохондриальной мембране с участием ферментов дыхательной цепи. Дыхательная цепь имеет чёткую структурную организацию, её компоненты формируют дыхательные комплексы, порядок расположения которых зависит от величины их редокс-потенциала (рис.5.1). Количество дыхательных цепей в отдельно взятой митохондрии из клеток разных тканей неодинаково: в печени – 5000, в сердце – около 20 000, следовательно, миокардиоциты отличаются более интенсивным дыханием, чем гепатоциты.
|
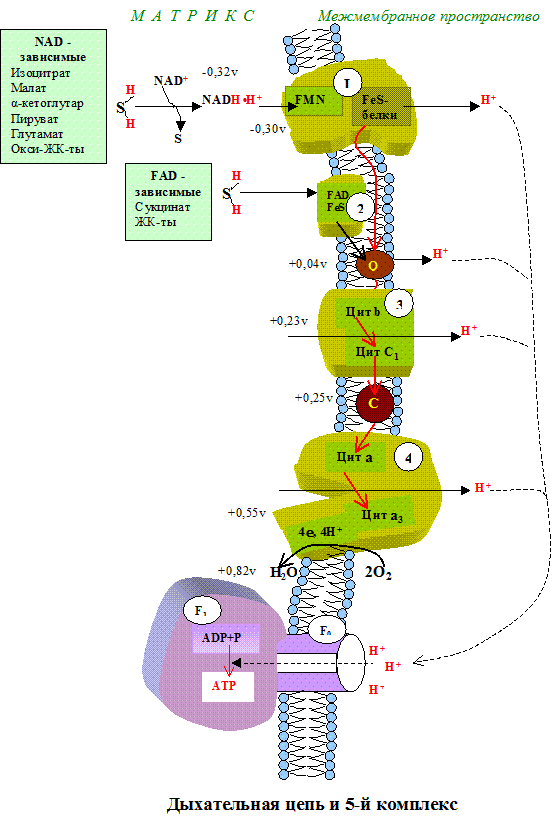
Прежде чем остановиться на характеристике каждого из компонентов дыхательной цепи, познакомимся с субстратами тканевого дыхания.
Субстраты тканевого дыхания подразделяются на 2 группы:
1. НАД-зависимые – субстраты цикла Кребса изоцитрат, α-кетоглутарат и малат. Это также пируват, гидроксибутират и β–гидрокси-ацил~КоА, глутамат и некоторые другие аминокислоты. Водород от НАД-зависимых субстратов c помощью НАД-зависимых дегидрогеназ передаётся на I-й комплекс дыхательной цепи.
2.ФАД-зависимые – сукцинат, глицерол-3-фосфат, ацил~КоА и некоторые другие. Водород от ФАД-зависимых субстратов передаётся на II-й комплекс дыхательной цепи.
При дегидрировании субстратов НАД-зависимыми дегидрогеназами образуется восстановленная форма НАД (НАДH∙H ).
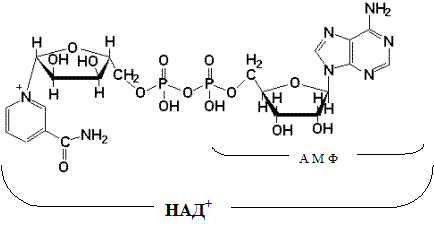
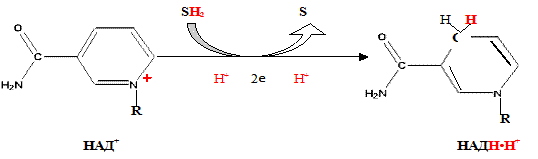
Указана окисленная форма кофермента НАД . Этот кофермент является динуклеотидом (никотинамид–аденин–динуклеотид): в состав одного нуклеотида входит витамин РР (никотинамид), другой представляет собой АМФ. Способность кофермента играть роль промежуточного переносчика водородов связана с наличием в его структуре витамина РР. В электронно-протонной форме процесс обратимого гидрирования-дегидрирования может быть представлен уравнением (R- остальная часть кофермента):
НАДH∙H может образовываться не только в митохондриях, но и в цитозоле клетки при протекании определённых процессов метаболизма. Однако цитоплазматический кофермент не может проникать в митохондрии. Водород восстановленного кофермента должен быть сначала перенесен на субстраты, которые могут проникать в митохондрии. Такими «Н2-переносящими субстратами» являются:
Оксалацетат → малат
Ацетоацетат → β-гидроксибутират
Дигидроксиацетон фосфат → глицерол-3-фосфат
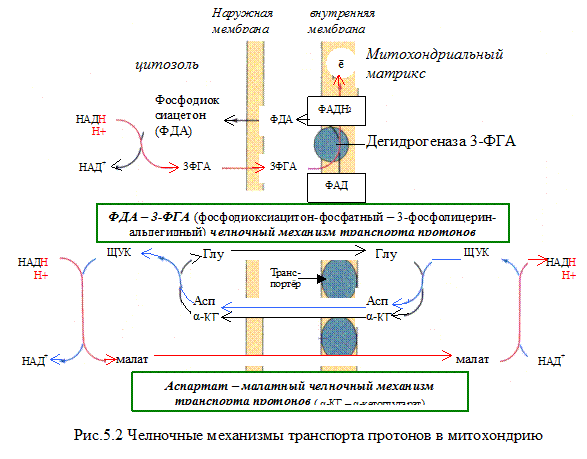
НАДH∙H затем окисляется 1-м комплексом дыхательной цепи. Рассмотрим работу этого комплекса.
I комплекс цепи тканевого дыхания – НАДH∙H -убихинон-оксидодуктаза.
Первый комплекс является самым большим в дыхательной цепи (представлен 23-30 субъединицами). Он катализирует перенос водорода от НАДH∙H на убихинон (рис. 5.1 и рис. 5.3). В его состав входят кофермент ФМН (флавинмононуклеотид) и железосерные белки, содержащие негеминовое железо. Функция этих белков заключается в разделении потока протонов и электронов: электроны переносятся от ФМН∙Н2 к внутренней поверхности внутренней мембраны митохндрий (обращенной к матриксу), а протоны – к внешней поверхности внутренней мембраны и затем высвобождаются в митохондриальный метрикс.
При транспорте протонов и электронов редокс-потенциал первого комплекса снижается на 0,38 v, что вполне достаточно для синтеза АТФ. Однако в самом комплексе АТФ не образуется, а высвобождающаяся в результате работы комплекса энергия аккумулируется (см. ниже образование электро-химического потенциала) и частично рассеивается в виде тепла.
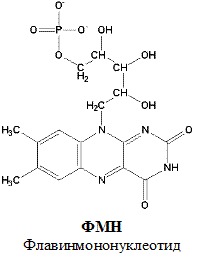
По своему строению ФМН – мононуклеотид, в котором азотистое основание представлено изоаллоксазиновым ядром рибофлавина, а пентозой является рибитол (иными словами, ФМН – это фосфорилированная форма витамина В2).
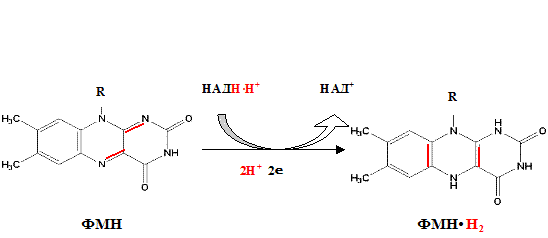
Функция ФМН заключается в акцепции 2 атомов водорода от НАДH∙H и передачи их железосерным белкам. Водород (2 электрона и 2 протона) присоединяется к атомам азота изоаллоксазинового кольца, при этом происходит внутримолекулярная перегруппировка двойных связей с образованием промежуточного семихинона – соединения свободнорадикальной природы (на схеме представлено суммарное уравнение реакции, где R – остальная часть молекулы)
II комплекс цепи тканевого дыхания –сукцинат-убихинон-оксидоредуктаза.
Этот комплекс имеет меньшую молекулярную массу и также содержит железосерные белки. Сукцинат-убихинон-оксидоредуктаза катализирует перенос водорода от сукцината на убихинон. В состав комплекса входит кофермент ФАД (флавин-аденин-динуклеотид) и фермент сукцинатдегидрогеназа, который является одновременно ферментом цикла Кребса. Ацил~SКоА, 3-фосфо-глицерат и диоксиацетон фосфат также являются ФАД-зависимыми субстратами тканевого дыхания и с помощью этого кофермента контактируют со вторым комплексом.
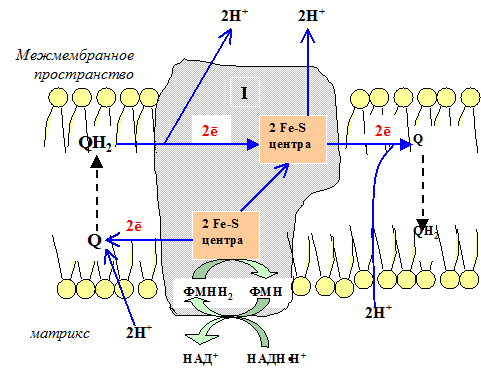
Рис. 5.3 Первый комплекс дыхательной цепи
Энергия включения водорода субстратов во II комплекс цепи тканевого дыхания рассеивается в основном в виде тепла, так как на этом участке цепи редокс-потенциал снижается незначительно и этой энергии для синтеза АТФ мало.
Процесс восстановления ФАД протекает аналогично таковому ФМН.
Кофермент Q или убихинон – гидрофобное соединение, является компонентом клеточных мембран, содержится в большой концентрации, относится к группе витаминов. относится к группе витаминов.
Убихинон (коэнзим Q). Убихинон – небольшая липофильная молекула, по химическому строению представляющая собой бензохинон с длинной боковой цепью (число изопреноидных единиц колеблется от 6 у бактерий до 10 у млекопитающих).
В дыхательной цепи коэнзим Q является своеобразным депо (пулом) водорода, который он получает от различных флавопротеинов. Липофильный характер молекулы убихинона обуславливает его способность свободно перемещаться в липидной фазе митохондриальной мембраны, перехватывая протоны и электроны не только от I и II комплексов дыхательной цепи, но и захватывая из митохондриального матрикса протоны. При этом убихинон восстанавливается с образованием промежуточного свободнорадикального продукта – семихинона .
Восстановленная форма убихинона – убихинол – передаёт протоны и электроны на III комплекс дыхательной цепи.
Цитохромоксидаза имеет высокую степень сродства к кислороду и может работать при его низких концентрациях.
аа3 – состоит из 6 субъединиц каждая из которых содержит гем и атом меди. 2 субъединицы составляют цитохром а, а остальные 4 относятся к цитохрому а3.
Между НАД и ФП, b-c, a-a3 имеет место max перепад ОВП. Эти пункты являются местом синтеза АТФ (местом фосфорилирования АДФ).
III комплекс цепи тканевого дыхания –убихинол-цитохром С-оксидоредуктаза. В состав III комплекса входят цитохромы b и с1, относящиеся к группе сложных белков хромопротеинов. Простетическая группа этих белков окрашена (chroma – краска) и близка по химическому строению к гему гемоглобина. Однако в противоположность гемоглобину и оксигемоглобину, в которых железо должно быть только в 2-х валентной форме, железо в цитохромах при работе дыхательной цепи переходит от двух- к трёхвалентному состоянию (и обратно).
Как видно из названия, III комплекс переносит электроны от убихинола на цитохром С. Вначале электроны поступают на окисленную форму цитохрома b (Fe3 ), который при этом восстанавливается (Fe2 ), затем восстановленный цитохром b передаёт электроны окисленной форме цитохрома с, который также восстанавливается и, в свою очередь, передаёт электроны цитохрому С.
|
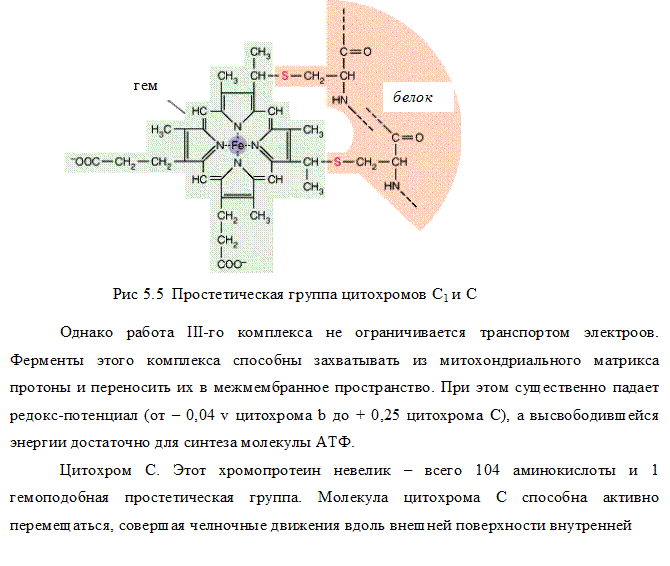
IV комплекс дыхательной цепи – цитохром С-оксидаза. Комплекс назван оксидазой из-за способности непосредственно взаимодействовать с кислородом. У млекопитающих этот крупный (~ 200 kD) трансмембранный белок состоит из 6-13 субъединиц, из которых некоторые кодируются митохондриальной ДНК. В состав IV комплекса входят 2 хромопротена – цитохром а и цитохром а3. В отличие от других цитохромов, цитохромы а и а3каждый содержат не только атом железа, но и атом меди. Медь в составе этих цитохромов при транспорте электронов также попеременно переходит в окисленное (Cu2 ) и восстановленное (Cu ) состояние.
Цитохром с-оксидаза катализирует одноэлектронное окисление 4-х восстановленных молекул цитохрома си при этом одновременно осуществляет полное (4-х электронное) восстановление молекулы кислорода:
 4 цитохрома с(Fe2 ) 4 H O2 4 цитохрома с(Fe3 ) H2O
4 цитохрома с(Fe2 ) 4 H O2 4 цитохрома с(Fe3 ) H2O
Протоны для образования молекул воды поступают из матрикса. Следует заметить, что эта реакция весьма сложна и протекает через промежуточные стадии образования свободных радикалов кислорода.
Окислительно-восстановительный потенциал IV комплекса является самым большим ( 0,57 v), его энергии вполне достаточно для синтеза 3-х молекул АТФ, однако большая часть этой энергии используется на «перекачивание» протонов из матрикса митохондрий в межмембранное пространство. В связи с активным транспортом протонов цитохром с-оксидаза получила название «протонного насоса».
Таким образом, тканевое дыхание представляет собой процесс транспорта электронов и протонов от НАД- или ФАД-зависимых субстратов на кислород, а также протонов, поставляемых матриксом митохондрий. При транспорте падает редокс-потенциал, что сопровождается высвобождением заключённой в субстратах тканевого дыхания энергии. Полное восстановление молекулярного кислорода воздуха в дыхательной цепи сопровождается образованием воды.
§
Ингибиторы дыхательной цепи являются ядами. Они в равной степени тормозят и дыхание, и фосфорилирование, соотношение Р/О при этом не изменяется. Наиболее популярными являются следующие ингибиторы: ротенон (растительный токсин, применяемый индейцами Амазонии при ловле рыбы, а также используемый в качестве инсектицида), амитал (аминобарбитал – производное барбитуровой кислоты), антимицин А (антибиотик) и цианиды. Ротенон и амитал тормозят перенос водорода от ФМН к убихинону; антимицин А тормозит транспорт электронов от цитохрома bк цитохрому с1; CN–, CO, H2S – ингибиторы цитохромоксидазы (рис.5. 8):

НАДН∙Н КоQ С О2
ротенон антимицин А СО, СN–
амитал H2S
Рис.5.8. Ингибиторы дыхательной цепи
Следует различать ингибиторы цепи тканевого дыхания и разобщители окислительного фосфорилирования.
3. Механизм генерации DmН , его компоненты, стехиометрия Н /е.
Движение электронов через электронтраспортную систему дыхательных ферентов сопровождается одновременно «перекачиваем» протонов из матрикса в межмебранное пространство. Эта передислокация протонов осуществляется I, III и IV комплексами дыхательной цепи. Английский биохимик Питер Митчелл предложил рассматривать внутреннюю мембрану митохондрий как конденсатор, который со стороны матрикса заряжается отрицательно (благодаря направленному току электронов), а со стороны межмембранного пространства – положительно. Следовательно, при тканевом дыхании совершается осмотическая работа по концентрации протонов в межмембранном пространстве и возникает разность электрических потенциалов, т.е. генерируется электро-химический (или протонный) потенциал:
Δ μН = Δ Ψ – (-)ΔрН
Электрохимический Мембранный Градиент рН
градиент рН потенциал
Градиент ΔрН имеет отрицательное значение, поскольку измерение проводится от внутренней поверхности мембраны, имеющей более отрицательную величину, к наружной. В целом Δ μН имеет положительное значение, равное 0,224v.
Внутренняя мембрана не может заряжаться сколь угодно долго. Если продолжить аналогию с конденсатором, то перезарядка приводит к «пробою», т.е. разрядке.Роль разряжающего устройства выполняет АТФ-синтаза – V комплекс дыхательной цепи.
Каким же образом генерирование протонного потенциала связано (сопряжено) с образованием АТФ?
§
Свободная энергия электрохимического протонного градиента митохондриальной мембраны используется для синтеза АТФ с помощью протон-переносящей АТФ-синтазы (она также называется протонным насосом, АТФ-азой, F1Fo-АТФ-азой, V комплексом дыхательной цепи).
АТФ-синтазаявляется самым крупным, по форме напоминающим гриб, структурным компонентом внутренней митохондриальной мембраны. АТФ-синтаза представлена 2 большими полиферметными белками – F1 (шляпка гриба)и Fo(ножка), каждый из которых, в свою очередь, состоит из нескольких неоднородных полипептидов.
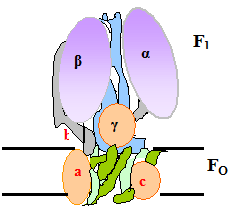
Рис. 5.6 . Модель грибовидного комплекса АТФ-синтазы. V комплекс.
Fо – не растворимый в воде трансмембранный белок, главной частью которого является канал для транслокации протонов. Fo означает «фактор О» (от слова олигомицин – антибиотик, продуцируемый Streptomyces). Олигомицин ингибирует транспорт протонов через «ножку» Fо, реагируя с единственным остатком глутаминовй кислоты на одной из субъединиц. Блокада транспорта протонов приводит к потере способности АТФ-азы синтезировать АТФ.
F1– водорастворимый периферический мембранный белок, состоящий из 5 субъединиц. Если отделить (например, мочевиной) F1 от Fо, то первый сможет только гидролизовать АТФ, но потеряет способность эту молекулу синтезировать.
Синтез АТФ катализирует β-субъединица белка F1. Этот синтез осуществляется спонтанно, т.е. без затраты энергии. Происходит это следующим образом (рис.5.7): F1 имеет 3 химически идентичных, но конформационно различно взаимодействующих αβ-протомеров: О (open) – с открытой конформацией, характеризующейся очень низкое сродство к лигандам и каталитически неактивной; L (loose) – легко связывающей лиганды и каталитически неактивной; T (tight) – жёстко связывающей лиганды и каталитически активной.
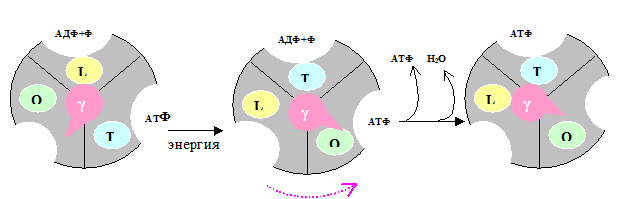
Рис. 5. 7. Работа «молекулярной турбины» – фактора F1.
В состав F1 входит γ-субъединица, способная вращаться подобно тому, как вращается турбина в воде («молекулярная турбина»). Вращение γ-субъдиницы обуславливает конформационные изменения в αβ-протомерах.
Связывание АДФ с неорганическим фосфатом происходит на участке L. После этого происходит вращение γ-субъединицы на 120˚ (на рис. против часовой стрелки). Это вращение требует затраты энергии, которая аккумулируется при генерации электро-химического потенциала и подпитывается протонным переносом через канал Fo. При вращении γ-субъединицы происходят конформационные изменения αβ-протомеров, в результате чего L-участок преобразуется в T, а T – в O. В Т-участке осуществляется синтез АТФ, а из О-участка (вследствие вращения «турбины») высвобождается АТФ.
Таким образом, энергия электро-химического потенциала требуется не для синтеза АТФ, а для удаления её из активного центра V комплекса. Эта энергия расходуется также на транспорт АТФ из матрикса через митохондриальные мембраны в цитоплазму клетки, а АДФ – в обратном направлении (на работу фермента АТФ-транслоказы). Львиная доля энергии протонного потенциала расходуется на транспорт ионов кальция. Доставка субстратов тканевого дыхания также осуществляется за счёт этой энергии.
Измерение «силы протонного потенциала» показало, что 1 молекула АТФ может образоваться за счёт окисления НАДН∙Н первым комплексом дыхательной цепи, 1 молекула АТФ – за счёт окисления убихинола третьим комплексом и 1 молекула – при окислении цитохрома счетвёртым комплексом. Именно в этих местах падение редокс-потенциала превышает 0,2 v, при таком перепаде освобождается энергия, вполне достаточная для синтеза молекулы АТФ (для синтеза АТФ необходимо затратить 42 кДж/моль). Таким образом, при окислении НАД-зависимых субстратов может образоваться 3 молекулы АТФ, а при окислении ФАД-зависимых – 2 молекулы АТФ (транспорт электронов начинается со второго комплекса дыхательной цепи).
Следует заметить, что ещё в 30-х годах академик В.А. Энгельгардт обнаружил, что при тканевом дыхании на 1 молекулу кислорода расходуется 3 молекулы фосфата, т.е. отношение Р/О=3 (при использовании НАД-зависиымых субстратов тканевого дыхания). Если в среду инкубации митохондрий добавлялся сукцинат, коэффициент Р/О=2. Эти показатели были названы В.А. Белицером и Е.Т. Цыбаковой коэффициентами фосфорилирования, а I, III и IV комплексы дыхательной цепи – пунктами сопряжения (связи между процессами окисления и фосфорилирования).
Согласно подсчётам известного российского биохимика В.П. Скулачёва, в сутки у человека может синтезироваться более 30 кг (!) АТФ: поскольку в сутки у человека образуется 400 мл воды (22 моля), а коэффициент Р/О=3 (66 молей АТФ), то умножая на молекулярную массу АТФ (507), получаем
707 х 66 = 33 462 (т.е.>33 кг)
§
Транспорт электронов (окисление НАДН∙Н и ФАДН2 кислородом) и окислительное фосфорилирование (синтез АТФ) в норме тесно связаны Это называется сопряжением окисления и фосфорилирования. В состоянии покоя, когда окислительное фосфорилирование минимально, электрохимический градиент внутренней митохондриальной мембраны достигает величины, при которой прекращается дальнейший перенос протонов. Тем самым ингибируется транспорт электронов.
Однако обнаружены вещества, например, 2,4-динитрофенол (ДНФ), которые разобщают эти процессы (ДНФ получил печальную известность как препарат для снижения массы тела). Присутствие во внутренней митохондриальной мембране агентов (ионофоров), увеличивающих их проницаемость для Н , разобщает окислительное фосфорилирование от процесса транспорта электронов, поскольку при этом нарушается генерирование электро-химического потенциала и, следовательно, синтез АТФ. ДНФ является липофильной слабой кислотой, поэтому легко проходит через мембрану (снаружи внутрь митохондрии), ослабляя естественный ток протонов, идущий в противоположном направлении.
Путь транспорта электронов, не связанный с синтезом АТФ, называется свободным, нефосфорилирующим, окислением. При свободном окислении энергия не аккумулируется, а высвобождается в виде тепла. Это имеет физиологическое значение при охлаждении организма.
Частичное разобщение окисления и фосфорилирования наблюдается при многих заболеваниях, так как митохондрии являются наиболее чувствительными клеточными органеллами к действию различных повреждающих факторов. Нарушение их структуры, приводящее к частичному или полному распаду внутренней митохондриальной мембраны, неизбежно будет способствовать обратному току протонов и нарушать энергопродукцию. Поэтому особое значение приобретает стабилизация митохондриальных мембран биоантиоксидантами (витаминами Е, А и аскорбатом) при любой патологии.
В ряде случаев некоторые пункты фосфорилирования могут «выключатся» – такое состояние называется разобщением окислительного фосфорилирования – и в этом случае P/O снижается: для НАД-зависимых субстратов – ниже 3; для ФАД-зависимых субстратов – ниже 2-ух.
И исходя из 1-го закона термодинамики в разобщенных митохондриях увеличивается теплообразование. (Это происходит за счет того, что та энергия электронов, которая должна была быть использована для синтеза АТФ в «выключенном» пункте фосфорилирования, рассеивается в виде тепла).
Процесс разобщения окислительного фосфорилирования лежит в основе лихорадки, вызванной бактериями, вирусами и другими агентами.
Разобщение резко усиливается при охлаждении организма.
Работа митохондрий при «выключенных» всех пунктах фосфорилирования называется сопряженной, в противном, выше описанном случае, разобщенной и дыхание при этом называется свободным.
В качестве разобщителей окислительного фосфорилирования выступают слабые гидрофобные кислоты (ЖК), тиреоидные гормоны, лекарства(дикумарин, динитрофенол).
ДЦ имеет механизмы шунтирования: сброс электронов и протонов по протяжению с НАД на цитохромы, или с НАД на межмембранные дегидрогеназы, на наружную мембрану и гладкую ЭПС (микросомальную цепь).
Такой перенос характерен для печени. При воздействии какого-либо блокатора, возникает блок в 1-ом комплексе ДЦ, происходит накопление НАД.Н2 и становится реальной угроза гипоксии, печень может погибнуть. Чтобы этого не произошло происходит сброс НАД.Н2 с митохондрий ДЦ на микросомальную ДЦ через цитохром b5.
§
Экспериментальные исследования показали, что добавление в ячейку полярографа с инкубируемыми митохондриями АДФ резко стимулировало потребление митохондриями кислорода. Накопление АДФ – регуляторный сигнал, вызывающий стимуляцию тканевого дыхания, т.е. усиление окисления субстратов тканевого дыхания. Это имеет большое физиологическое значение, так как увеличение концентрации АДФ в клетке является свидетельством низкой концентрации АТФ (соотношение адениловых нуклеотидов в клетке постоянно), следовательно, указывает на энергетическое её голодание. Отсюда вытекает и необходимость большего потребления субстратов. Помимо регуляторной роли соотношения АДФ/АТФ, в контроле скорости фосфорилирования участвуют НАД-зависимые коферменты: высокое соотношение НАДН∙Н /НАД , так же как и увеличение соотношения АТФ/АДФ будет снижать интенсивность тканевого дыхания.
Способность генерации энергии присуща всем тканям но для того, чтобы они все зароботали, необходимо сильное охлаждение организма.
Поэтому в организме есть ткань, которая обеспечивает термогенез в обычных условиях. Это бурая жировая ткань (БЖТ). Ее особенно много у новорожденных (от затылка до крестца, вдоль всей спины). У взрослого она локализуется в в определенных местах: между лопаток, в паху. Ярко выраженной способностью к теплопродукции обладает бурая жировая ткань новорождённых (а её много в организме зимнеспящих животных), которая отличается от типичной жировой ткани тем, что содержит много триацилглицеролов. Кроме того, в митохондриях этой ткани так много цитохромов, что она приобретает бурый цвет. Митохондрии бурой жировой ткани содержат разобщающий белок термогенин (димер 32 кД), который действует подобно каналу, контролирующему проводимость протонов во внутренней митохондриальной мембране. Поток протонов через этот канал ингибируется физиологическими концентрациями пуриновых нуклеотидов (АДФ, АТФ, ГДФ и др), но это ингибирование может быть устранено свободными жирными кислотами. Компоненты этой системы подчиняются гормональному контролю, в частности, норадреналину, который через систему цАМФ активирует гормончувствительную липазу, расщепляющую триацилглицеролы бурого жира с освобождением свободных жирных кислот.
БЖТ имеет большое количество митохондрий, т. к. митохондрии содержат цитохромы, то это и придает этой ткани бурый цвет. Особенностью митохондрий БЖТ является отсутствие АТФ-азы, DmН генерируется в обычном режиме, но нет инструмента (аденилаттранслаказы), трансформирующего DmН —> АТФ. Вместо нее есть белок термогенин, который шунтирует DmН с наружной мембраны во внутрь и одновременно способствует рассеиванию энергии DmН в виде тепла, так и происходит подогрев тканей. БЖТ охватывает крупные кровеносные сосуды и согревает кровь, а потом эта кровь согревает перефирические участки тела.
Это и есть несократительный термогенез.
Механизм сократительного термогенеза связан с окислительным фосфорилированием.
При охлаждении организма, активизируется симпатическая нервная система, в овет на это происходит выброс адреналина, под действием которого идет гликолиз, через аденилатциклазный механизм. Образующиеся при этом ЖК разобщают окислительное фосфорилирование и (согласно 1-му закону термодинамики) теплопродукция увеличивается. Именно поэтому, после принятия жирной пищи наступает состояние температурного комфорта.
6. Основополагающая роль энергетического обмена. Пути утилизации DmН и АТФ. Прикладные аспекты биоэнергетики.
Энергетический обмен играет ведущую роль в жизнедеятельности организмов, т. к. все функции организма энергозависимы. Систему механизмов, обеспечивающих стабильный уровень субстратов энергообмена называют энергетическим гомеостазом.
Одним из механизмов поддержания постоянного уровня АТФ в клетке, является наличие мегамитохондрий, которое дает большое преимущество.
Если один участок клетки плохо снабжается кислородом, то при помощи мегамитохондрий энергия DmН транспортируется в этот участок и восполняет недостаток АТФ.
§
Тканевое дыхание – один из процессов диссимиляции, по сути это и есть биологическое окисление в тканях и клетках организма. В организме существует 3 пути потребления и утилизации кислорода:
1 путь – 90-95% O2 идет на митохондриальное окисление.
2 путь – 5-10% идет на микросомальное окисление (в печени при поступлении больших количеств токсинов – 40%).
3 путь – перекисное окисление (2-5%).
Микросомальная дыхательная цепь.
Микросомы (микрочастицы) – это замкнутые мембранные пузырьки (везикулы), образуемые из гладкой ЭПС при гомогенизации клетки. Как таковых микросом не существует.
Микросомальное окисление – это окисление, протекающее на гладкой ЭПС нормальной неразрушенной клетки.
Наиболее интенсивно микросомальное окисление протекает в печени и надпочечниках, а также в местах контакта с внешней средой, в коже, почках, легких, селезенке.
ЭПС – 2-й слой мембран, ассоциированных с 3-мя основными классами ферментов:
1) оксидоредуктазы;
2) трансферазы;
3) гидролазы.
Главная функция этих ферментов – реакции детоксикации.
Микросомальное окисление осуществляется с помощью одноименной ДЦ, которая представляет собой систему переносчиков протонов и электронов с НАД или НАДФ на кислород.
Существует 2 варианта микросомальной ДЦ:
1) НАДФ —-> ФП —> b5 —> p450 —> O2
2) НАД —-> ФП —-> b5—–
Цитохром b5 одной цепи может передавать свои электроны на цитохром b5 другой цепи, а также на цитохром p450.
Микросомальное окисление можно записать и так:
RH НАД (НАДФ).H2 O2 —> ROH НАД (НАДФ) HOH
схема
FP – флавопротеид, включающий ФАД и Fe-белок, содержащий негеминовое железо.
P450 – восстановленный CO-комплекс, который имеет max поглощения при длине волны = 450 нм.
Многие гидрофобные вещества организма обладают токсичностью, за счет того, что растворяются в клеточных мембранах и тем самым разрушают их.
Задачей организма является перевод этих гидрофобных соединений в гидрофильные, которые легче выводятся почками. Это осуществляется микросомальным окислением.
Таким образом, основная роль микросомальной ДЦ заключается в осуществлении реакций синтеза с участием кислорода (в схеме видно образование фенола из бензола (гидроксилирование)).
Для связывания второго атома кислорода необходим косубстрат, каковым является аскорбат (Vit C), кторый также отдает 2H на синтез H2O. Для обеспечения реакций детоксикации необходимо большое количество Vit C в составе косубстрата: реакции детоксикации протекают по механизму гидроксилирование гетероциклических и алифатических соединений (ксенобиотики), поступающие из вне.
Реакции детоксикации могут привести к снижению концентрации токсических веществ или может возникнуть летальный синтез.
Роль микросомального окисления состоит в биосинтезе Vit D, кортикостероидов, коллагена, тирозина, катехоламинов.
– Реакции деалкилирования – отщепление алкильной группы;
– реакции окисления спиртов, альдегидов, кетонов, нитросоединений;
– реакции разрыва кольца ароматических соединений;
– реакции восстановления, когда идет сброс протов;
– реакции десатурации (перевод насыщенных ЖК в ненасыщенные).
В 70-е годы было показано, что микросомальная и митохондриальная дыхательные цепи взаимодействуют друг с другом через цитохром b5.
В условиях интоксикации (этанол, барбитураты) происходит ингибирование 1 комплекса митохондриальной ДЦ.
НАД —> ФП -/-> Q —> b —> c1 —> c —> aa3 —> ½ O2
Окисление НАД.Н2 не происходит и он накапливается. В межмембранном пространстве имеется цитохром b5, который принимает электроны с НАД.Н М/Х ДЦ и перебрасывает их на микросомальную ДЦ и тем самым угроза энергетического голода устраняется.
Таким образом ц. b5 – фермент, компонент микросомальной ДЦ, который обеспечивает межмембранный митохондриально-микросомальный перенос электронов.
Сходства и различие митохондриальной и микросомальной дыхательных цепей.
1. Сходства: а) они имеют одинаковые начало и конец и одинаковую суммарную разность потенциалов (а значит одинаковый градиент энергии в начале и конце);
б) имеют одинаковые переносчики: НАД, ФП, цитохромы.
2. Различия: а) по локализации;
б) микросомальная ДЦ короче и электроны на последнем переносчике М/С цепи более энергизированы и спосбны активировать кислород;
в) будучи активным кислород способен внедряться в структуру многих молекул, т. е. используется с «пластическими» целями (ФЕН—> ТИР). В то время как в М/Х ДЦ кислород – всего лишь конечный акцептор электронов и используется в энергетических целях;
г) в процессе переноса электронов в М/Х ДЦ их энергия депонируется в форме АТФ. В М/С ДЦ – депонирование энергии ни в каком виде не происходит;
д) М/С окисление – современная интерпретация теории БахаЭнслера. М /Х окисление – современный вариант теории Палладина-Виланда.
Перекисное окисление. Механизм образования активных форм кислорода. Роль перекисных процессов в норме и при патологии. Общее представление о ПОЛ (НЭЖК → R· → диеновые коньюгаты → гидроперекиси → МДА). Способы оценки активности ПОЛ.
Перекисное окисление и антиоксидантная защита.
Еще Мечников, изучая фагацитоз утверждал, что фагоцитарное действие лейкоцитов осуществляется за счет перекисных процессов.
Перекисное окисление – это третий путь утилизации вдыхаемого кислорода (от 2 до 5%).
Кислород сам по себе является парамагнитным элементом (это было установлено методом молекулярных орбиталей) т. к. имеет на внешнем слое 2 неспаренных электрона.
——— _ _ .
——— O2; O2 e —> O2, т. е. в реакциях перекисного
— окисления происходит одноэлектронное восстановление
кислорода.
——— _.
——— O2 – супероксидный ион-радикал, более активная форма
— кислорода.
Возможна еще одна активная форма кислорода:
——— _
——— O2 – синглетный кислород.
_. _
O2 и O2 – инициируют образование большого количества радикалов, по цепному механизму:
_. _.
O2 H —> HO2 – гидропероксидный радикал
_. _.
HO2 H O2 —-> H2O2 O2 .
H2O2 Fe2 —-> Fe3 OH- OH (пероксидный радикал).
_.
O2 Fe3 —> O2 Fe2
В процессе взаимодействия этих радикалов с веществом поражаются наиболее уязвимые места клеток: ненасыщенные ЖК фосфолипидов мембран, они «выжигаются» в результате чего мембрана делается более ригидной и следовательно изменяется ответная реакция клетки.
В нормальных условиях перекисное окисление регулирует агрегатное состояние мембран, лежит в основе тканевой адаптации. (Это играет роль в стрессовых ситуациях, когда клетка т. о. защищается от избытка гормонов).
При всех видах патологии активность перекисных процессов возрастает, и является инструментом повреждения мембраны. В ней образуются мощные ионные каналы, через которые входят ионы Na , K и др. и содержимое клетки как бы вываливается и она гибнет.
OH. – радикал взаимодействует с ДНК и РНК, вызывая возникновение генных мутаций и провоцируя канцерогенез.
Перекисные процессы инициируются в структуре нуклеиновых кислот.
§
Клетки имеют мощную антиоксидантную защиту, состоящую из двух уровней:
1) ферментативная (происходит восстановление продуктов перекисного окисления и их ликвидация с помощью ферментов):
а) супероксиддисмутаза – сложный фермент, встречаются Mg, Zn, Fe, Cu – содержащие формы в разных тканях. Его активность повышается при любых формах активации перекисных процессов. Этот фермент за рубежом выделяется в чистом виде и эффективно используется в лучевой терапии. Действие СОД направлено на супероксид ион:
СОД
О2* О2* 2Н ———– Н2О2 О2
б) каталаза (её субстратом является Н2О2) особенно активна в эритроцитах, которые специализируются на переносе кислорода:
2Н2О2 ——— 2Н2О О2
в) пероксидаза – наиболее активна глутатион – пероксидаза
г) глутатион – редуктаза – является непосредственным защитником эритроцитов, в частности предохраняют от образования МеHb, который не способен к транспорту кислорода, что ведёт к гипоксии. МеHb образуется при приёме избытка нитратов, аспирина, сульфаниламидов.
К ферментативной антиоксидантной защите относятся ферменты, генерирующие восстановительную форму НАД * Н и НАДФ * Н (такую систему имеют все клетки, но особенно мозг и миокард), а также ферменты, поддерживающие восстановительную форму металлов.
2) неферментативная: сюда относится ряд легко окисляющихся веществ, обладающих меньшей активностью, чем естественные метаболиты:
– хинон
– убихинон (Ко – Q)
– витамины Е и А (являются компонентами мембран и блокируют перекисные процессы)
– витамин С
Между этими тремя витаминами существует взаимосвязь: витамин С обеспечивает восстановительную форму витамина Е, а для поддержания восстановительной формы витамина С нужен витамин А. В настоящее время существует мощный препарат антиоксидантной защиты, представляющий собой комплекс трёх витаминов (Vit C = 2 г, Vit E = 500000 E, Vit A 140000 – 170000 Е). Витамин А довольно токсичен, поэтому в качестве замены используется В – каротин.
Также к антиоксидантам относятся Vit F, кортикостероиды, гистидин, аргинин, билирубин и растительные пигменты.
| Заведующий кафедрой биологической химии, д.м.н., проф. | Грицук А. И. | ___________ |
21.10.2006
Министерство здравоохранения Республики Беларусь
УО «Гомельский государственный медицинский университет»
Кафедра биологической химии
Обсуждено на заседании кафедры (МК или ЦУНМС)
Протокол № _________________200__года
ЛЕКЦИЯ
по биологической химии
наименование дисциплины
для студентов _2__ курса лечебного факультета
Тема Биохимия крови-1. Основы регуляции КОС.
Время 90 мин.
Учебные и воспитательные цели:
Дать представление:
1. О кислотно-основном состоянии (КОС). О принципах регуляции (изоосмолярность, электронейтральность, постоянство pH). О механизмах регуляции КОС (физико-химических, физиологических). О характеристике и механизме действия буферов. О функции экскреторных систем. О способах оценки КОС крови (pH, pCO2, BЕ, HCO3–, ионов Na , K Cl– плазмы и эритроцитов, pH мочи).
2. О белках крови (методах фракционирования, осаждения, электрофореза). О характеристиках основных фракций – альбумине и глобулинаъ. О белковом коэффициенте. О функциональной классификации белков плазмы крови. О белках острой фазы. Об остаточном азоте, его диагностическом значение.
3. Об особенностях метаболизма эритроцита. Об антиоксидантной системе. О роли GSH, ПФЦ, гликолиза, изоцитрат- и малатдегидрогеназ, о роли генетических дефектов — гемолитические анемии.
ЛИТЕРАТУРА
МАТЕРИАЛЬНОЕ ОБЕСПЕЧЕНИЕ
1. Мультимедийная презентация.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
| № п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
| 1. | Определение КОС. Принципы регуляции КОС. Изоосмолярность, электронейтральность, постоянство pH. Механизмы регуляции КОС – физико-химический (забуферивание, разбавление, фиксация H в слабодиссоциируемые соединения). Характеристика, и механизм действия буферов. Физиологические механизмы- функции экскреторных систем. Способы оценки КОС крови (pH, pCO2, BЕ, HCO3–, ионов Na , K Cl– плазмы и эритроцитов, pH мочи). | |
| 2. | Белки крови (методы фракционирования, осаждения, электрофорез). Характеристика основных фракций – альбумин и глобулины. Белковый коэффициент. Функциональная классификация белков плазмы крови. Белки острой фазы. Остаточный азот, его диагностическое значение. | |
| 3. | Особенности метаболизма эритроцита. Антиоксидантная система. Роль GSH, ПФЦ, гликолиза, изоцитрат и малатдегидрогеназ, генетические дефекты — гемолитические анемии. |
Всего 90 мин
| Заведующий кафедрой биологической химии, д.м.н., проф. | Грицук А. И. | ___________ |
21.10.2006
Министерство здравоохранения Республики Беларусь
УО «Гомельский государственный медицинский университет»
Кафедра биологической химии
Обсуждено на заседании кафедры (МК или ЦУНМС)
Протокол № _________________200__года
ЛЕКЦИЯ
по биологической химии
наименование дисциплины
для студентов _2__ курса лечебного факультета
Тема Биохимия крови-2. Обмен гемоглобина.
Время 90 мин.
Учебные и воспитательные цели:
Дать представление:
1. Об особенностях метаболизма эритроцитов крови. О шунте Раппопорта. О роли 2,3-ДФГК. О биосинтезе, строении гема. О роли железа, характеристике белковой части Hb. О динамике изменений типов Hb в онтогенезе. О распаде Hb в норме и при патологии (уровень билирубина). О дифференциальной диагностике желтух (гемолитической, обтурационной, паренхиматозной).
2. Об обмене железа, всасывании, транспорте, депонировании.
3. Об особенностях обмена лейкоцитов. О фагоцитозе. О биохимии тромбоцитов.
ЛИТЕРАТУРА
МАТЕРИАЛЬНОЕ ОБЕСПЕЧЕНИЕ
1. Мультимедийная презентация.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
| № п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
| 1. | Особенности метаболизма эритроцитов крови. Шунт Раппопорта. Роль 2,3-ДФГК. Биосинтез, строение гема. Роль железа, характеристика белковой части Hb. Динамика изменений видов Hb в онтогенезе. Распад Hb в норме и при патологии (уровень билирубина). Дифференциальная диагностика желтух (гемолитическая, обтурационная, паренхиматозная). | |
| 2. | Обмен железа, всасывание, транспорт, депонирование. | |
| 3. | Особенности обмена лейкоцитов. Биохимия фагоцитоза. Биохимия тромбоцитов. |
Всего 90 мин
| Заведующий кафедрой биологической химии, д.м.н., проф. | Грицук А. И. | ___________ |
21.10.2006
Переваривание углеводов
Основными пищевыми углеводами являются крахмал и дисахариды. Для взрослых суточная потребность в углеводах составляет 400-600 г, для детей 12 г/кг.
В ротовой полости происходит частичное расщепление крахмала под действием α – амилазы слюны, которая расщепляет в крахмале внутренние 1,4 –α – гликозидные связи с образованием декстринов и очень небольшого количества мальтозы.
В желудке отсутствуют ферменты переваривания углеводов, а резко кислая среда ингибирует активность амилазы, поэтому переваривание продолжается только внутри пищевого комка.
Основное переваривание углеводов происходит в тонком кишечнике под действием ферментов поджелудочной железы и слизистой кишечника. Наиболее активным ферментом, осуществлющим полостное переваривание углеводов, является
lang=”ru-RU” lang=”ru-RU”>панкреатическая α – амилаза, расщепляющая 1,4 –α -гликозидные связи до точек ветвления крахмала. Ответвления (1,6 –α -гликозидные связи) расщепляется дополнительным ферментом –
Ферменты кишечника представлены α- гликозидазами и β – гликозидазами, осуществляющими пристеночное пищеварение углеводов. Основными кишечными ферментами являются:
- мальтаза (расщепляет 1,4 –альфа-гликозидные связи в дисахариде мальтозе),
- изомальтаза (расщепляет 1,6 – гликозидные связи в крахмале),
- сахараза (расщепляет 1, 2- α, β гликозидные связи в дисахариде сахарозе),
- лактаза (расщепляет 1,4- β-гликозидные связи в дисахариде лактозе),
- гетерогалактозидаза – расщепляет гликозидные связи смешанных олигосахаридов,
β – галактозидаза с оптимумом рН 5.
Под действием перечисленных ферментов полисахариды и дисахариды расщепляются до моносахаридов.





