

Клетка – элементарная биологическая система. клеточная теория т.шванна и м.шлейдена, история, её основные положения. современное состояние клеточной теории. значение клеточной теории.
Конец XIX века – возникновение цитологии
1665 – англ. Роберт Гук, рассматривая срез пробки, увидел целлюлозные оболочки и ввел термин «клетка».
1838 – 1839 – М.Шлейден и Т. Шванн предложили клеточную теорию.
Клеточная теория
1. Все организмы состоят из клеток.
2. Все клетки развиваются по единому плану.
3. Свойства многоклеточного организма сводятся к арифметической сумме свойств тех клеток, которые его слагают.
Шлейден предложил считать ядро наиболее постоянной структурой клетки. Многие положения оказались неверными (положение 3). Клетка стала изучаться. Клеточная теория оказала большое влияние на биологию и медицину.
1858 – Рудольф Вирхов(РОБЕРТ РЕМАРК) опубликовал свой труд. Если существует живая клетка, то она произошла от клетки. « Каждая клетка от клетки». Применил свои положения теории в клеточной патологии.
Современная клеточная теория
1. Жизнь существует только в форме клеток.
2. в основе непрерывности жизни лежит клетка.
3. Принцип комплиментарности (связь между структурой и функцией).
КЛЕТКА — ЭЛЕМЕНТАРНАЯ ЕДИНИЦА ЖИВОГО
Клетка представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка, таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль элементарной структурной, функциональной и генетической единицы.
Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия.
Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой.
КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория сформулирована немецким исследователем, зоологом Т. Шванном (1839). Поскольку при создании этой теории Шванн широко пользовался работами ботаника М. Шлейдена, последнего по праву считают соавтором клеточной теории. Исходя из предположения о схожести (гомологичности) растительных и животных клеток, доказываемой одинаковым механизмом их возникновения, Шванн обобщил многочисленные данные в виде теории, согласно которой клетки являются структурной и функциональной основой живых существ.
В конце XIX столетия немецкий патолог Р. Вирхов на основе новых фактов пересмотрел клеточную теорию. Ему принадлежит вывод о том, что клетка может возникнуть лишь из предсуществующей клетки. Им также создана вызвавшая критику концепция «клеточного государства», согласно которой многоклеточный организм состоит из относительно самостоятельных единиц (клеток), поставленных в своей жизнедеятельности в тесную зависимость друг от друга.
Клеточная теория в современном виде включает три главных положения.
Первое положение соотносит клетку с живой природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые (например, вирусы) формы она ни принимала, в ее структурном, функциональном и генетическом отношении обеспечивается в конечном итоге только клеткой. Выдающаяся роль клетки как первоисточника жизни обусловливается тем, что именно она является биологической единицей, с помощью которой происходит извлечение из внешней среды, превращение и использование организмами энергии и веществ. Непосредственно в клетке сохраняется и используется биологическая информация.
Второе положение указывает, что в настоящих условиях единственным способом возникновения новых клеток является деление предсуществующих клеток. В обосновании клеточной природы жизни на Земле тезису о единообразии путей возникновения клеток принадлежит особая роль. Именно этот тезис был использован М. Шлейденом и Т. Шванном для обоснования представления о гомологии разных типов клеток. Современная биология расширила круг доказательств этому. Независимо от индивидуальных структурно-функциональных особенностей все клетки одинаковым образом: а) хранят биологическую информацию, б) редуплицируют генетический материал с целью его передачи в ряду поколений, в) используют информацию для осуществления своих функций на основе синтеза белка, г) хранят и переносят энергию, д) превращают энергию в работу, е) регулируют обмен веществ.
Третье положение клеточной теории соотносит клетку с многоклеточными организмами, для которых характерен принцип целостности и системной организации. Для системы свойственно наличие новых качеств благодаря взаимному влиянию и взаимодействию единиц, составляющих эту систему. Структурно-функциональными единицами многоклеточных существ являются клетки. Вместе с тем многоклеточный организм характеризуется рядом особых свойств, которые нельзя свести к свойствам и качествам отдельных клеток. В третьем положении клеточной теории мы встречаемся с проблемой соотношения части и целого.
Системный подход как научное направление используется в биологических исследованиях с начала прошлого столетия. Системный характер организации и функционирования свойствен не только организму, но и другим главным биологическим образованиям — геному, клетке, популяции, биогеоценозу, биосфере.
9.Биологическая мембрана, молекулярная организация и функции. Транспорт веществ через мембрану (модели транспорта).
Клетка – система мембран, отграничивающих участки внутриклеточного пространства. Мембраны участвуют в разных процессах. Мембраны нервных клеток – генерация нервного импульса, мембраны ЖКТ – всасывание и переваривание пищи, клеточные мембраны скелетных мышц и клеток миокарда – расслабление и сокращение, мембраны клеток органов чувств – преобразование одного вида раздражения в другой. Белки природных мембран плохо растворимы в воде, образуют комплексы с липидами. Функции: рецепторная, структурная, транспортная, каталитическая (большинство белков – ферменты – иммуноглобулины – белки с наибольшей активностью). Жидкостно-мозаическап модель строения мембраны (бислой липидов, белки – периферические, погруженные, интегральные). Транспорт веществ не всегда происходит путем диффузии или градиента. Существуют транспортные белки.
АТФ—АДФ Ф
Активный транспорт – перенос веществ через мембрану с затратой АТФ и при участии транспортных белков. Активный сопряженный транспорт (одни и те же белки – несколько веществ). Могут быть 2 периферических белка, могут идти вещества по каналу, 2-3 переносчика, транспорт может быть несопряженным. Бывает экзцитоз (пиноцитоз и фагоцитоз). Существование обменной диффузии (при помощи градиента концентрации),
В эукариотических клетках эндоплазматическая мембрана составляет единое целое с внутриклеточными мембранами (цитоплазматические мембраны). Цитоплазматические мембраны образуют канал со сложной сетью переплетающихся каналов и пузырьков (1959 Картер) каналы ЭПР – временные коммуникационные системы, участвующие в перемещении пузырьков из клетки наружу. Мембраны гЭПР имеют трубчатые структуры (более молодой эволюционный тип). Биологическая роль: гидролиз фосфолипидов, синтез стероидных гормонов, синтез липидов и т.д. как и плазматическая, цитоплазматическая мембрана переходит в ядерную и комплекс Гольджи(1898 ит. Гольджи) главная особенность – отсутствие рибосом. Это плоскопараллельные цистерны, пузырьки. Комплекс специализирован для синтеза белков. Собственные клеточные белки, секреторные белки синтезируются в большинстве клеток. Большинство белков сложные, имеют углеводную и белковую часть – гликопротеины и протеогликаны. Комплекс Гольджи участвует в синтезе сложных сахаров из простых моносахаридов, которые затем связываются с белками, пришедшими в комплекс, возникают гликопротеины, образовавшись, они отшнуровываются, происходит экзоцитоз. В комплексе Гольджи образуются лизосомы. Комплекс Гольджи поставляет необходимый материал для образования борозд дробления в животной клетке.
Высокая упорядоченность внутреннего содержимого эукариотиче-ской клетки достигается путем компартментации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава. Компартментация (рис. 2.3) способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии).

Рис. 2.3. Компартментация объема клетки с помощью мембран:
1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4—транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 — гранула секрета

Рис. 2.4. Молекулярная организация биологической мембраны:
1 — бимолекулярный слой липидов, 2 — белки
Предложено несколько схем взаимоотношения в мембране основных химических компонентов — белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время принята точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные — находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях (рис. 2.4).
Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаимодействуют друг с другом.
10.Ядро. Строение и функции.
В клетке выделяют ядро и цитоплазму. Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети.
Ядерная оболочка пронизана порожу диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Подсчитано, что у низших позвоночных животных в эритробластах, где интенсивно образуется и накапливается гемоглобин, на 1 мкм2 ядерной оболочки приходится около 30 пор. В зрелых эритроцитах названных животных, сохраняющих ядра, на 1 мк»г оболочки остается до пяти пор, т.е. в 6 раз меньше.
В области перового комплекса начинается так называемая плотная пластинка — белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре.
Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации — гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК (см. 3.4.3.2).
Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомальных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15 и 21—22 пары) — ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называются вторичными перетяжками. С помощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент.
Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки (см. 3.5.2.2).
§
В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки.
Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3 нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов см. разд. 2.3.8.
Включениями (рис. 2.5) называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значения и специальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества — переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии.
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез.
Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети (см. рис. 2.3). Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой.
Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.
Рибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма —с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока).
Пластинчатый комплекс Голъджи образован совокупностью диктиосом числом от нескольких десятков (обычно около 20) до нескольких сотен и даже тысяч на клетку.
Диктиосома (рис. 2.6, А) представлена стопкой из 3—12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий.в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки.
Митохондрии (рис. 2.6, Б) — это структуры округлой или палочко-видной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5—10 мкм. В большинстве животных клеток количество митохондрий колеблется от 150 до 1500, однако в женских половых клетках их число достигает нескольких сотен тысяч. В сперматозоидах нередко присутствует одна гигантская митохондрия, спирально закрученная вокруг осевой части жгутика. Одна разветвленная митохондрия обнаружена в клетке такого паразита человека, как трипаносома.
Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20—40 нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген.
В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала — этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки (чем подтверждается симбиотическая гипотеза происхождения митохондрий; см. § 1.5). Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме.
Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата —АТФ). В целом этот процесс называется окислительным (расформированием. В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая).
Лизосомы (рис. 2.6, В) представляют собой пузырьки диаметром обычно 0,2—0,4 мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом — внутриклеточное переваривание оазличных химических соединений и структур.
Первичными лизосомами (диаметр 100 нм) называют неактивные органеллы, вторичными — органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы (фаголизосомы) и аутолизосомы (цитолизосомы). В первых (рис. 2.6, Г) переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называют остаточными тельцами (телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал.
Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1—1,5 мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает 70—100.
К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки (рис. 2.6, Д) — трубчатые образования различной длины с внешним диаметром 24 нм, шириной просвета 15 нм и толщиной стенки около 5 нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов.
Микрофиламентами (рис. 2.6, Е) называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы.
По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм — промежуточные филстенты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию.
Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации.
Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Центриолъ (под электронным микроскопом) имеет вид «полого» цилиндра диаметром около 150 нм и длиной 300—500 нм. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза.
Эукариотическая клетка имеет клеточный скелет (цитоскелет) из внутриклеточных волокон (Кольцов) – начало ХХ века, в конце 1970 вновь открыт. Эта структура позволяет клетке иметь свою форму, иногда изменяя ее. Цитоплазма находится в движении. Цитоскелет участвует с процессе переноса органоидов, участвует в регенерации клеток.
Митохондрии – сложные образования с двойной мембраной(0,2-0,7мкм) и разной формой. Внутренняя мембрана имеет кристы. Наружная мембрана проницаема практически для всех химических веществ, внутренняя – только активный транспорт. Между мембранами – матрикс. Митохондрии располагаются там, где необходима энергия. Митохондрии имеют систему рибосом, молекулу ДНК. Возможно возникновение мутаций (более66 заболеваний). Как правило, они связаны с недостаточной энергией АТФ, часто связаны с сердечно-сосудистой недостаточностью, патологиями. Количество митохондрий разное (в клетке трипаносомы- 1 митохондрия). Количество зависит от возраста, функции, активности ткани (печень – более1000).
Лизосомы – тельца, окруженные элементарной мембраной. Содержат 60 ферментов( 40 лизосомальных, гидролитических). Внутри лизосомы – нейтральная среда. Активизируются низкими значениями рН, выходя в цитоплазму (самопереваривание). Мембраны лизосом защищают цитоплазму и клетку от разрушения. Образуются в комплексе Гольджи (внутриклеточный желудок, могут перерабатывать отработавшие свое структуры клетки). Есть 4 вида. 1-первичные, 2-4 – вторичные. С помощью эндоцитоза в клетку попадает вещество. Первичная лизосома (запасающая гранула) с набором ферментов, поглощает вещество и образуется пищеварительная вакуоль (при полном переваривании расщепление идет до низкомолекулярных соединений). Непереваренные остатки остаются в остаточных тельцах, которые могут накапливаться (лизосомные болезни накопления). Остаточные тельца, накапливающиеся в эмбриональном периоде, приводят к гаргалеизму, уродствам, мукополисахаридозам. Аутофагирующие лизосомы уничтожают собственные структуры клетки( ненужные структуры). Могут содержать митохондрии, части комплекса Гольджи. Часто образуются при голодании. Могут возникать при воздействии других клеток (эритроциты).
12.Поток информации, энергии и вещества в клетке.
Жизнедеятельность клетки как единицы биологической активности обеспечивается совокупностью взаимосвязанных, приуроченных к определенным внутриклеточным структурам, упорядоченных во времени и пространстве обменных (метаболических) процессов. Эти процессы образуют три потока: информации, энергии и веществ.
Благодаря наличию потока информации клетка на основе многовекового эволюционного опыта предков приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, а также передает в ряду поколений.
В потоке информации участвуют ядро (конкретно ДНК хромосом), макромолекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). На завершающем этапе этого потока полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуры и используются в качестве катализаторов или структурных белков (рис. 2.7). Кроме основного по объему заключенной информации ядерного генома в эукариотических клетках функционируют также геномы митохондрий, а в зеленых растениях — и хлоропластов.
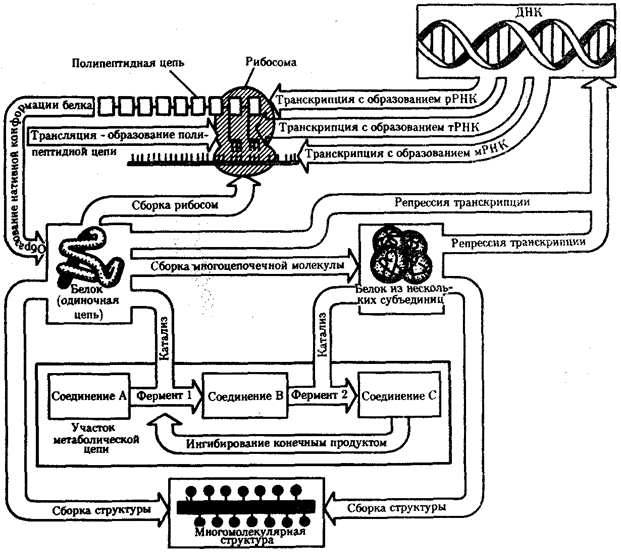
Рис. 2.7. Поток биологической информации в клетке
§
1. Ген – это часть молекулы ДНК, которая является функциональной единицей наследственной информации.
1. Ген занимает определенный участок в хромосоме – локус.
2. Внутри гена может происходить перекомбинация.
3. ДНК, входящая в состав гена способна к репарации.
4. Существуют гены: структурные, регуляторные и т.д.
5. Расположение триплетов комплиментарно аминокислотам (мутации со сдвигом рамки считывания).
6. Генотип, будучи дискретным (состоящим из отдельных генов) функционирует как единое целое.
7. Генетический код универсален.
8. Генетический код вырожден (для многих аминокислот существует более одного кодона – сайта)
9. Гены располагаются в хромосоме в линейном порядке и образуют группу сцепления. Число групп сцепления соответствует гаплоидному набору хромосом (23 у человека или 24 с оговоркой на пол – х и у хромосомы).
Структура белков определяется набором и порядком расположения аминокислот в их пептидных цепях. Именно эта последовательность аминокислот в пептидах зашифрована в молекулах ДНК с помощью генетического кода.
Свойства генетического кода:
- Триплетность- каждая аминокислота шифруется тремя стоящими рядом нуклеотидами.
- Вырожденность- многие аминокислоты шифруются несколькими триплетами.
- Специфичность– каждый триплет способен кодировать только одну определенную аминокислоту.
- Универсальность– полное соответствие кода у различных видов живых организмов.
- Непрерывность– последовательности нуклеотидов считываются триплет за триплетом без пропусков.
- Неперекрываемость кодонов– соседние триплеты не перекрывают друг друга.
20.Рибосомный цикл синтеза белка (инициация, элонгация, терминация). Посттрансляционные преобразования белков.
Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК, но непосредственного участия в жизнеобеспечении клетки она не принимает. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют РНК. Процесс взаимодействия мРНК и тРНК, обеспечивающий трансляцию информации с языка нуклеотидов на язык аминокислот, осуществляется на рибосомах. Рибосомные РНК являются не только структурным компонентом рибосом, но и обеспечивают связывание их с определенной нуклеотидной последовательностью мРНК. Этим устанавливается начало и рамка считывания при образовании пептидной цепи. Кроме того, они обеспечивают взаимодействие рибосомы и тРНК. В рибосомах имеется 2 бороздки. Одна из них удерживает растущую полипептидную цепь, другая-мРНК. Кроме того, в рибосомах выделяют 2 участка, связывающих тРНК. В аминоацильном, А-участке размещается аминоацил-тРНК, несущая определенную аминокислоту. В пептидильном, П-участке располагается обычно тРНК. Образование А- и П-участков обеспечивается обеими субчастицами рибосомы. В ходе трансляции можно выделить 3 фазы: инициацию, элонгацию, терминацию.
Фаза инициации заключается в объединении двух находящихся порознь до этого в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК.
Фаза элонгации или удлинении пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней амк. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил –тРНК очередного кодона, находящегося в А-участке, комплементарное вз-е между антикодоном и кодоном. Фаза терминаци или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ, УГА), когда тот входит в зону входит А-участка рибосомы. При этом к последней амк в пептидной цепи присоединяется вода и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы.
Посттрансляционное преобразование белков.Синтезированные в ходе трансляции пептидные цепи на основе своей первичной структуры приобретают вторичную и третичную, а многие четвертичную организацию, образуемую несколькими пептидными цепями. В зависимости от функций, выполняемых белками, их аминокислотные последовательности могут претерпевать различные преобразования, формируя функционально активные молекулы белка. Многие мембранные белки синтезируются в виде пре-белков, имеющих на N-конце лидерную последовательность, которая обеспечивает им узнавание мембраны. Секреторные белки также имеют на N-конце лидерную последовательность, которая обеспечивает их транспорт через мембрану. Некоторые белки сразу после трансляции несут дополнительные аминокислотные про-последовательности, определяющие стабильность предшественников активных белков. При созревании белка они удаляются, обеспечивая переход неактивного пробелка в активный белок. Формируя третичную и четвертичную организацию в ходе посттрансляционных преобразований, белки приобретают способность активно функционировать, включаясь в определенные клеточные структуры и осуществляя ферментативные и другие функции.
Взаимосвязь между геном и признаком. Гипотеза «один ген – один фермент», ее современная трактовка: «один ген – одна полипептидная цепь»
Ген – участок молекулы ДНК, который несет информацию о структуре полипептидной цепи или макромолекулы. Гены одной хромосомы располагаются линейно, образую группу сцепления. ДНК в хромосоме выполняет разные функции. Гены имеют маленький размер, хотя состоят из тысяч пар нуклеотидов. Наличие гена устанавливается по проявлению признака гена (конечному продукту).
Для Менделя, ген – только символ, удобный для определения закона наследования. Связь между геном и признаком (продуктом) была открыта при изучении брожения в безвоздушной среде – 1902 г Гаррод. Он изучал родословные больных алкаптонурией, пришел к выводу, что болезнь – результат нарушения обмена азота, при этом. Вместо мочевины образуется темное вещество. При содействии Бэтса в 1908 году высказано предположение, что болезнь возникает у рецессивных гомозигот, у которых не хватает какой-то ферментативной реакции, что приводит к накоплению и выведению субстрата, который в норме должен был расщепиться. В крови людей содержится гомогентизиновая кислота, но в норме она расщепляется оксидазой гомогентизиновой кислоты до малеинацетата, затем до воды и углекислого газа. У больных нет оксидазы, поэтому происходит накопление кислоты и вывод ее с мочой.
Так же наследуется альбинизм, хотя встречается гораздо чаще. При этом заболевании отсутствует фермент, осуществляющий превращение тирозина в меланин.
До 1940 года мнение ученых разделялось, но теории не было.1940 год – Бидл и Татумпредложили гипотезу: 1 ген – 1 фермент. Эта гипотеза сыграла важную роль – ученые стали рассматривать конечные продукты. Оказалось, что гипотеза имеет ограничения, т.к. все ферменты – белки, но не все белки – ферменты. Как правило, белки являются олигомерами – т.е. существуют в четвертичной структуре. Пример, капсула табачной мозаики имеет более 1200 полипептидов. В настоящее время наиболее приемлемой считают гипотезу «Один ген – один полипептид». Под термином ген надо понимать функциональную единицу наследственности, по химической природе являющуюся полинуклеотидом и определяющую возможность синтеза полипептидной цепи.
Ген как единица изменчивости. Генные мутации и их классификация. Причины и механизмы возникновения генных мутаций. Последствия генных мутаций.
Ген представляет собой элементарную единицу наследственного материала.Генные мутации связаны с изменением внутренней структуры генов, что превращает одни аллели в другие.
Изменения структуры ДНК, образующей ген, можно разделить на 3 группы:
§
Процесс образования яйцеклеток в яичниках овогенез, оогенез. Сперматозоиды образуются в семенниках, процесс носит название сперматогенеза. Те и другие клетки образуются по-разному, но есть некоторые общие черты.
Сперматогенез. Морфологически семенник состоит из множества семенных канальцев. Дольчатое строение. Между семенными канальцами – клетки Лейдинга (начинают работу в 12-14 лет) синтезируют тестостерон – развитие вторичных половых признаков. Семенник очень рано становится эндокринным органом, под влиянием андрогенов происходит формирование мужских половых органов. Семенной каналец имеет зоны:
-размножения, -роста,- созревания и формирования.
Существуют одноименные периоды роста. Зона размножения в наружной части семенника. Клетки округлые, цитоплазмы много, ядро большое – сперматогонии. Они размножатся митозом, и семенник увеличивается в размерах до полового созревания, после – делятся только стволовые клетки. Запас клеток не уменьшается и семенник тоже не уменьшается. В зоне размножения 2n2c.следующая фаза – роста. Увеличивается размер ядра, цитоплазмы, идет репликация ДНК (интерфаза 1), клетки – сперматоциты первого порядка 2n4c. Эти клетки вступают в зону формирования и созревания у семенных канальцев. Мейоз состоит из 2 митотических делений, после первого деления n2c, после второго – nc.
Овогенез (яичники). Половые железы закладываются на 2м месяце эмбрионального развития. У человека очень рано закладывается желточный мешок (функция формирования первичных половых клеток, обеспечение питательными веществами). Половые клетки (первичные) мигрируют в развивающуюся половую железу, а желточный мешок дегенерирует. В эмбриогенезе яичники не активны. Формирование женских половых клеток пассивное. Первичные половые клетки – овогонии, они делятся. Формируются овоциты первого порядка. Период деления оканчивается к 7му месяцу эмбриогенеза – 7000000 первичных клеток. 400-500 созревают в течение жизни, остальные невостребованы. Развитие яйцеклеток у человека блокируется в профазе первого мейотического деления (на стадии диплотены). С наступлением половой зрелости овоцит увеличивается в размере, растет и размер желтка. Накапливаются пигменты, происходят биохимические и морфологические изменения. Каждый овоцит окружается мелкими фолликулярными клетками, созревающими в фолликуле. Яйцеклетка, созревая, приближается к периферии. Фолликулярная жидкость окружает её на всех этапах. Фолликул разрывается. Яйцеклетка попадает в брюшную полость. Затем в воронку яйцевода. Продолжение мейоза в 2/3 яйцевода в результате контакта яйцеклетки со сперматозоидом.
При мейозе идет распределение хромосом. В результате 4 ядра. Происходит конъюгация хромосом (за счет высоко повторяющихся последовательностей ДНК в 1ген). Каждое из 4х ядер при гаметогенезе получает только 1 хроматиду из пары. В результате мейоза при сперматогенезе из каждого спермацита первого порядка получаются 4 хроматиды и формируются 4 сперматозоида. Из одного овоцита первого порядка образуются 2 ядра с гаплоидным набором хромосом. Одно из них, с большим количеством цитоплазмы (т.к. при цитокинезе разделение идет неравномерно) и другое – редукционное (направляющее) тельце. При последующем делении образуются яйцеклетка и направляющее тельце. При овогенезе из каждого овоцита формируется 1 яйцеклетка и 3 направляющих тельца, которые дегенерируют и исчезают. В яйцеклетке есть все необходимые запасы питательных веществ. Мейоз– способ распределения хромосом, генов, обеспечивающий их независимую и случайную рекомбинацию. При овогенезе служит для перераспределения цитоплазмы между клетками. Кроссинговер – способ, осуществляющий сближение и перераспределение генов отдельных гомологичных хромосом.
С 212-218
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
· Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
· Фаза лептотены или лептонемы — конденсация ДНК с образованием хромосом в виде тонких нитей.
· Зиготена или зигонема — конъюгация (соединение) гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами.
· Пахитена или пахинема — кроссинговер (перекрест), обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой.
· Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой.
· Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; гомологичные хромосомы остаются соединёнными между собой.
· Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
· Анафаза I — микротрубочки сокращаются, биваленты делятся и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
· Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
· Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления.
· Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
· Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
· Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений).
Морфология половых клеток.
Зрелые половые клетки содержат гаплоидный набор хромосом. Созревающие -диплоидный. Имеют ядро, цитоплазму, клеточные органеллы. Несмотря на это, строение мужских и женских половых клеток неодинаково. Это объясняется различными функциями. Функции сперматозоида – оплодотворение (стимуляция дальнейшего развития яйцеклетки), обеспечение генетической информацией мужского организма. Все сперматозоиды имеют жгутики, подвижны, небольшого размера (50-90мкм у человека). Состоят из головки, шейки, средней части и хвостика. Головка -5мкм, шейка – 5. головка сперматозоида почти полностью занята ядром, цитоплазмы мало, она в жидкокристаллическом состоянии (защита от вредных явлений – ионизирующего излучения). Находится по периферии ядра. На конце головки – акросома с видоизмененным комплексом Гольджи. Ферменты: гиалуронидаза, муциназа. В плазматической мембране – проакрозин, который превращается в акрозин, проходя по половым путям самки (происходит отщепление ингибитора). Функция акрозина – отщепление фолликулярных клеток, отщепление блестящей оболочки. Шейка содержит пару центриолей. Микротрубочки одной из них удлиняются, образуется основная нить хвостика. В шейке много митохондрий, расположенных по спирали. Органеллы движения – жгутики, способны к биению только при смешивании с секретом. Предстательной железы при семяизвержении. При нарушении функций предстательной железы – мужская стерильность.
Яйцеклетка .Функции: передает зародышу половину его будущего хромосомного набора; во время оплодотворения яйцеклетка приносит гораздо больше цитоплазмы; яйцеклетка снабжает зародыш пищевыми запасами до начала его собственного питания.
Размеры яйцеклеток много больше размеров сперматозоидов(130-150 мкм у человека). В зрелой яйцеклетке запасаются все материалы, которые обеспечивают начальные стадии развития зародыша. Если сперматозоид, созревая, старается избавиться от цитоплазмы, яйцеклетка, наоборот, стремится увеличить ее количество. Есть рибосомы, рРНК, т-РНК, морфогенетические факторы. Многие белки синтезируются в печени, жировом теле, а затем транспортируются в яйцеклетку. Яйцеклетка имеет плазматическую мембрану. Во время оплодотворения плазматическая мембрана контролирует поступление многих ионов (например, натрия). К ней прилегает желточная оболочка (гликопротеины – специфическое прикрепление сперматозоида своего вида к соответствующей яйцеклетке), часто прозрачна, яйцеклетка окружена слоем клеток лучистого яйца – фолликулярными питающими клетками. Для оплодотворения сперматозоид должен пройти сквозь все оболочки. Наследственный материал, приносимый яйцеклеткой и сперматозоидом по размеру одинаков
32.Оплодотворение, его фазы, биологическая сущность. Партеногенез. Типы определения пола.
Зрелая яйцеклетка может быть оплодотворена на протяжении небольшого отрезка времени(12-24 часа после овуляции у человека, у рыб и амфибий – через несколько минут после откладки). После овуляции яйцеклетка претерпевает серьезные клеточные изменения. Для оплодотворения очень важен срок жизни. Совокупность изменений яйцеклетки после овуляции – старение яйцеклетки, через 24 часа – перезрелая яйцеклетка – оплодотворение невозможно. Если происходит оплодотворение в данном случае, то невозможна имплантация. Возможны аборты. Ежемесячно созревает только 1 яйцеклетка.
Жизнеспособность сперматозоидов зависит от многих факторов. В щелочной среде они активны, но быстро погибают в кислой среде. Путь от влагалища до 2/3 яйцевода составляет 2-3 часа. В верхней трети влагалища происходит подщелачивание среды (т. к. кислотность высокая) из-за семенной жидкости. В матку проникают только сперматозоиды. Они сохраняют большую подвижность при пониженной температуре. Сперма собирается и замораживается в жидком азоте. Жидко кристаллическая цитоплазма не позволяет образовываться большим кристаллам льда (использование в племенном деле, банки спермы людей). В процессе оплодотворения выделяют 3 этапа.
1) активация яйцеклетки
2) проникновение сперматозоида в яйцеклетку
3) слияние ядер.
В ходе оплодотворения сперматозоид преодолевает все оболочки яйцеклетки. Как только происходит контакт сперматозоида с яйцеклеткой – завершается мейоз яйцеклетки, в т.ч. она выделяет фертилизин. Сперматозоид выделяет антифертилизин – происходит приклеивание. В это время акрозин разрушает оболочки, выделяются ферменты. У человека сперматозоид проникает по-разному: либо только головка, либо шейка и головка. После контакта сперматозоида и яйцеклетки у моноспермных в течение 1-3 минут происходит кортикальная реакция – образуется оболочка оплодотворения. Если несколько сперматозоидов – гибель клетки. Ядро сперматозоида набухает, разрыхляется, образуется мужской пронуклеус. Ядро яйцеклетки превращается в женский пронуклеус. Идет репликация ДНК. Пронуклеусы идут к центру, ядерные оболочки исчезают, происходит слияние пронуклеусов (кариогамия), образуется зигота(2n4c). Через сутки начинается дробление. Возможно развитие эмбриона без участия мужской гаметы (механическое, тепловое, химическое воздействие) – партеногенез. Гиногенез – развитие из яйцеклетки. При гиногенезе сперматозоид проникает и погибает (только женские организмы, т.к. только женский геном). Андрогенез – развитие из сперматозоида. При нм ядро яйцеклетки погибает (только мужской геном). Очень быстро такой геном перерождается в злокачественную опухоль хорионэпителиому.
Типы определения пола.
– прогамный. Пол будущего организма определяется в ходе гаметогенеза у родительских особей.
сингамный. Пол будущего организма определяется в момент слияния половых клеток.
– эпигамный. Пол будущего организма определяется в процессе онтогенеза. У человека имеет место переопределение пола (при патологии) – хотя истинного нет
Партеногенез- развитие без оплодотворения. В случае естественного партеногенеза развитие идет на основе цитоплазмы и пронуклеуса яйцеклетки. Естественный партеногенез- явление редкое, обнаружен у пчел, ос, ряда чешуекрылых. Искусственный партеногенез возможен, по-видимому, у всех животных. На тутовом шелкопряде было показано, что с помощью искусственного партеногенеза можно регулировать соотношение мужского и женского пола в популяции. В активированных яйцах используется информация только женского пронуклеуса – гиногенез.При искусственном партеногенезе можно удалить женский пронуклеус, тогда развитие осуществится за счет мужских пронуклеусов – андрогенез
33.Предмет, задачи, методы генетики. История развития генетики. Роль отечественных ученых (Н. И. Вавилов, Н. К. Кольцов, А. С. Серебровский, С. С. Четвериков) в развитии генетики.
Генетика человека – основа медицинской генетики. Человек – удобный генетический объект. У человека лучше, чем у других видов изучены биохимические, иммунологические, физиологические и другие реакции, а эти признаки детерминированы генами.
98-99% заболеваний возникают в результате поражения генетического материала.
1-2% – травмы и ожоги.
Генетика человека – наука о наследственно обусловленных различиях людей и о нарушениях генетического материала.
40-50 из 1000 детей с наследственной или врожденной патологией в РФ.
40%смертности, инвалидности из-за наследственных заболеваний.
30-40 коек в больницах – дети с врожденной или наследственной патологией.
Более половины таких детей погибают после рождения или получают хронические заболевания, остальные постоянно нуждаются в наблюдении врачей.
11-16% больных в педиатрических отделениях имеют генетические заболевания. Наследственные заболевания лечатся.
8,5% детей умирают от заболеваний, связанных с генными мутациями.
2,5% умирают от хромосомных (т.к. их мало рождается).
31% умирают от «отягощенности наследственных признаков».
42% умирают от наследственных заболеваний.
17% – от неизвестной патологии.
Большой процент больных наследственными патологиями в гематологических клиниках (до 40%)
16% – в детской нефрологии.
50% – детская слепота и инвалидность.
50% – нарушения слуха и инвалидность
наследование гипертонии у 60%(мультифакториальные, моногенные).
В РФ созданы генетические центры(90). В США их – 6, и они очень специализированы. Там проводят биохимические исследования.
Тяжелые формы наследственной патологии – 1-2%
1966 год – американский ученый Майкьюсик выявил 1487 наследственных признаков.
2005 год – 6678
Из них – аутосомно-доминантных – 4458 Аутосомно-рецессивных – 1730
Х-сцепленных – 421 У-сцепленных – 19 Митохондриальных – 66.
Составлен каталог наследственных признаков человека, который постоянно пополняется. Среди Х-сцепленных – ломкость Х хромосомы (снижение умственных способностей). Синтез митохондриальных белков под двойным контролем. Нарушается ДНК в митохондрии, нарушается синтез АТФ – сердечно-сосудистые заболевания. Митохондриальные = материнские болезни. Большое значение имеют болезни пероксисом (образованы округлым телами с простой мембраной, в которых есть кристаллический белок Д=0,3-0,1 мкм, возникают из ЭПР). Они ферментативно катализируют расщепление перекиси кислорода на воду и кислород под действием фермента – каталазы. Наследственные нарушения пероксисомного окисления лежат в основе заболеваний ЦНС, поражения органов зрения, печени, аномалии скелета. Известно 11 таких заболеваний. Особая группа болезней – лизосомальные болезни накопления – 15 заболеваний. Накопление токсичных продуктов в клетке. Число наследственных заболеваний возрастает, т.к. окружающая среда ухудшается, улучшается диагностика, происходит выхаживание больных детей, появляются чужеродные токсические химические соединения, происходит рост знаний.
В медицинском ВУЗе существуют обязательные программы медицинской генетики.
§
Признак – единица морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности организмов (клеток), т.е. отдельное качество или свойство, по которому они отличаются друг от друга.
Генотип — это генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе — кариотипе.
Генотип (от ген и тип), совокупность всех генов, локализованных в хромосомах данного организма.
Фенотип (Phenotype) — присущая индивидууму совокупность всех признаков и свойств, которые сформировались в процессе его индивидуального развития.
Фенотип – совокупность всех признаков организма, сформировавшаяся во взаимодействии генотипа с окружающей средой.
Гомозиготность, состояние наследственного аппарата организма, при котором гомологичные хромосомы имеют одну и ту же форму данного гена.
Гетерозиготность, присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена.
Гемизиготность (от греч hemi- — полу- и zygotós — соединённый вместе), состояние, связанное с тем, что у организма один или несколько генов не парные, т. е. не имеют аллельных партнёров. (В сцепленном с полом наследовании, Хr или ХR — r – дальтонзим)
35.Закономерности наследования при моногибридном скрещивании.
Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков.
1 закон Менделя: при скрещивании двух гомозиготных организмов, различающихся друг от друга по одной паре альтернативных признаков в первом поколении наблюдается единообразие по генотипу и по фенотипу. (фиброматоз десен — А, здоровые десны — а, ребенок в любом случае болен)
2 закон менделя: при скрещивании 2х гетерозиготных организмов, различающихся по одной паре альтернативных признаков (гибриды F1) в их потомстве (гибриды F2) наблюдает расщепление по фенотипу 3:1, по генотипу 1:2:1
Полное доминирование — явление, при котором один из аллельных генов имеет преобладающее значение и проявляется как в гетерозиготном, так и в гомозиготном состоянии.
36.Дигибридное и полигибридное скрещивание. Закон независимого комбинирования генов и его цитологические основы. Общая формула расщепления при независимом наследовании.
Дигибридное скрещивание – скрещивание форм, различающихся по двум парам изучаемых признаков
Полигибридное скрещивание –скрещивание форм, отличающихся по многим признакам.
Закон независимого наследования признаков:
При скрещивании гомозиготных особей, которые отличаются двумя и большим количеством пар альтернативных признаков, во втором гибридном поколении (при инбридинге гибридов 1 поколения) фиксируют независимое наследование по каждой паре признаков и появляются особи, с новыми сочетаниями признаков, не свойственных родительским и прародительским формам (закон независимого распределения, или III закон Менделя) (Карие глаза — B, голубые — b, правша — A, левша — a). Расщепление в отношении (3:1)n , а по фенотипу 9:3:3:1. Задача в альбоме.
Очевидно, этому закону должны подчиняться в первую очередь неаллельные гены, располагающиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования признаков объясняется закономерностями поведения негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами разные пары, или биваленты, которые в метафазе I мейоза случайно выстраиваются в плоскости экватора веретена деления. Затем в анафазе I мейоза гомологи каждой пары расходятся к разным полюсам веретена независимо от других пар. В результате у каждого из полюсов возникают случайные сочетания отцовских и материнских хромосом в гаплоидном наборе (см. рис. 3.75). Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неал-лельных генов.
Разнообразие типов гамет, образуемых организмом, определяется степенью его гетерозиготности и выражается формулой 2n, где n — число локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды F1 образуют четыре типа гамет с одинаковой вероятностью. Реализация всех возможных встреч этих гамет при оплодотворении приводит к появлению в F2 четырех фенотипических групп потомков в соотношении 9:3:3:1. Анализ потомков F2 по каждой паре альтернативных признаков в отдельности выявляет расщепление в соотношении 3:1.
37.Множественные аллели. Наследование групп крови человека системы АВО.
Множественный аллелизм — различные состояния (три и более) одного и того же локуса хромосом, возникшие в результате мутаций.
Присутствие в генофонде вида одновременно различных аллелей гена называют множественным аллелизмом. Примером этому служат разные варианты окраски глаз у плодовой мухи: белая, вишневая, красная, абрикосовая, эозиновая,— обусловленные различными аллелями соответствующего гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA, IB, I0). Два аллеля имеет ген, обусловливающий резус-принадлежность. Более ста аллелей насчитывают гены α- и β-полипептидов гемоглобина.
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков. Задачи в альбоме.
38.Взаимодействие неаллельных генов: комплементарность, эпистаз, полимерия, модифицирующее действие.
Комплементарность — такой тип взаимодействия, когда 2 неаллельных гена, попадая в генотип в доминирующем состоянии, совместно определяют появление нового признака, который каждый из них по отдельности не детерминирует.(R- розовидный гребень, P – гороховидный, rp – листовидный, RP – ореховидный)
Если присутствует один из пары – проявляется он.
Примером служат группы крови у человека.
Комплементарность может быть доминантная и рецессивная.
Для того чтобы человек имел нормальный слух, необходимо чтобы работали, согласовано многие гены, и доминантные и рецессивные. Если, хотя бы по одному гену он будет гомозиготен по рецессиву – слух будет ослаблен.
Эпистаз — маскирование генов одной аллельной пары генами другой.
Эпистаз (от греч. epi – над stasis — препятствие) — взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллелъного гена.
Ген, подавляющий фенотипические проявления другого, называется эпистатичным; ген, чья активность изменена или подавлена, называется гипостатичным.
Это обусловлено тем, что ферменты катализируют разные процессы клетки, Когда на одном метаболическом пути действуют несколько генов. Действие их должно быть согласовано во времени.
Механизм: если В выключится, он замаскирует действие С

В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе.
Полимерия — явление, когда различные неаллельные гены могут оказывать однозначное действие на один и тот же признак, усиливая его проявление.
Наследование признаков при полимерном взаимодействии генов. В том случае, когда сложный признак определяется несколькими парами генов в генотипе и их взаимодействие сводится к накоплению эффекта действия определенных аллелей этих генов, в потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4) (см. рис. 3.80). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 24 = 16 типов гамет, получается потомство, 1/256 которого имеет максимальную пигментацию кожи, 1/256 — минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака. В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента.
В некоторых случаях доминантные и рецессивные аллели полигенов могут обеспечивать развитие разных вариантов признаков. Например, у растения пастушьей сумки два гена одинаково влияют на определение формы стручочка. Их доминантные аллели образуют одну, а рецессивные — другую форму стручочков. При скрещивании двух дигетерозигот по этим генам (рис. 6.16) в потомстве наблюдается расщепление 15:1, где 15/16 потомков имеют от 1 до 4 доминантных аллелей, а 1/16, не имеет доминантных аллелей в генотипе.
Если гены располагаются, каждый в своем отдельном локусе, но их взаимодействие проявляется в одном и том же направлении – это полигены. Один ген проявляет признак незначительно. Полигены дополняют друг друга и оказывают мощное действие – возникает полигенная система – т.е. система является результатом действия одинаково направленных генов. Гены подвергаются значительному влиянию главных генов, которых более 50. полигенных систем известно множество.
При сахарном диабете наблюдается умственная отсталость.
Рост, уровень интеллекта – определяются полигенными системами
Модифицирующее действие. Гены модификаторы сами по себе не определяют какой- то признак, но могут усиливать или ослаблять действие основных генов, вызывая таким образом изменение фенотипа. В качестве примера обычно приводится наследование пегости у собак и лошадей. Числового расщепления никогда не даётся, так как характер наследования больше напоминает полигенное наследование количественных признаков.
1919 год Бриджес ввел термин ген-модификатор. Теоретически любой ген может взаимодействовать с другими генами, а значит, и проявлять модифицирующее действие, но некоторые гены – модификаторы в большей степени. Они часто не имеют собственного признака, но способны усиливать или ослаблять проявление признака, контролируемого другим геном. В формировании признака кроме основных генов проявляют свое действие и модифицирующие гены.
Брахидактилия – может быть резкая или незначительная. Помимо основного гена, есть еще модификатор, который усиливает эффект.
Окраска млекопитающих – белая, черная модификаторы.
39.Хромосомная теория наследственности. Сцепление генов. Группы сцепления. Кроссинговер как механизм, определяющий нарушения сцепления генов.
§
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
Гены расположены в хромосоме в линейной последовательности.
Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом (рис. 3.72). Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах.

Рис. 3.72. Кроссинговер как источник генетического разнообразия гамет:
I — оплодотворение родительских гамет а и б с образованием зиготы в; II — гаметогенез в организме, развившемся из зиготы в; г — кроссинговер, происходящий между гомологами в профазе I; д — клетки, образовавшиеся после 1-го мейотического деления; е, ж — клетки, образовавшиеся после 2-го деления мейоза (е — некроссоверные гаметы с исходными родительскими хромосомами; ж — кроссоверные гаметы с перекомбинацией наследственного материала в гомологичных хромосомах)
Понятно, что кроссинговер как механизм рекомбинации эффективен лишь в том случае, когда соответствующие гены отцовской и материнской хромосом представлены разными аллелями. Абсолютно идентичные группы сцепления при кроссинговере не дают новых сочетаний аллелей.
Кроссинговер происходит не только в предшественницах половых клеток при мейозе. Он наблюдается также в соматических клетках при митозе. Соматический кроссинговер описан у дрозофилы, у некоторых видов плесеней. Он осуществляется в ходе митоза между гомологичными хромосомами, однако его частота в 10 000 раз меньше частоты мейотического кроссинговера, от механизма которого он ничем не отличается. В результате митотического кроссинговера появляются клоны соматических клеток, различающихся по содержанию в них аллелей отдельных генов. Если в генотипе зиготы данный ген представлен двумя разными аллелями, то в результате соматического кроссинговера могут появиться клетки с одинаковыми либо отцовскими, либо материнскими аллелями данного гена (рис. 3.73).

Рис. 3.73. Кроссинговер в соматических клетках:
1 — соматическая клетка, в гомологичных хромосомах которой ген А представлен двумя разными аллелями (А и а); 2 — кроссинговер; 3 — результат обмена соответствующими участками между гомологичяыми хромосомами; 4 — расположение гомологов в плоскости экватора веретена деления в метафазе митоза (два варианта); 5 — образование дочерних клеток; 6 — образование гетерозитотиых по гену А клеток, сходных с материнской клеткой по набору аллелей (Аа); 7 — образование гомозиготных по гену А клеток, отличающихся от материнской клетки по набору аллелей (АА или аа)
§
Никакие признаки не наследуются. Признаки развиваются на основе взаимодействия генотипа и среды. Наследуется только генотип, т.е. комплекс генов, который определяет норму биологической реакции организма, изменяющую проявление и выраженность признаков в разных условиях среды. Таким образом, организм реагирует на свойства внешней среды. Иногда один и тот же ген в зависимости от генотипа и от условий внешней среды по-разному проявляет признак или меняет полноту выраженности.
Степень проявления фенотипа – экспрессивность. Образно ее можно сравнить со степенью тяжести болезни в клинической практике. Экспрессивность подчиняется законам распределения Гаусса (некоторые в малом или среднем количестве). В основе изменчивости экспрессивности лежат и генетические факторы, и факторы внешней среды. Экспрессивность – очень важный показатель фенотипического проявления гена. Количественно ее степень определяют, используя статистический показатель.
Экспрессивность также является показателем, характеризующим фенотипическое проявление наследственной информации. Она характеризует степень выраженности признака и, с одной стороны, зависит от дозы соответствующего аллеля гена при моногенном наследовании или от суммарной дозы доминантных аллелей генов при полигенном наследовании, а с другой — от факторов среды. Примером служит интенсивность красной окраски цветков ночной красавицы, убывающая в ряду генотипов АА, Аа, аа, или интенсивность пигментации кожи у человека, увеличивающаяся при возрастании числа доминантных аллелей в системе полигенов от 0 до 8 (см. рис. 3.80). Влияние средовых факторов на экспрессивность признака демонстрируется усилением степени пигментации кожи у человека при ультрафиолетовом облучении, когда появляется загар, или увеличением густоты шерсти у некоторых животных в зависимости от изменения температурного режима в разные сезоны года.
Генетический признак может даже не проявляться в некоторых случаях. Если ген есть в генотипе, но он вовсе не проявляется – он пенетрирован. (русский ученый Тимофеев-Рисовский 1927 год). Пенетрантность – количество особей (%), проявляющих в фенотипе данный ген, по отношению к количеству особей, у которых этот признак мог бы проявиться. Пенетрантность свойственна проявлению многих генов. Важен принцип – «все или ничего» – либо проявляется, любо нет.
– наследственный панкреатит – 80%
– вывих бедра – 25%
– пороки развития глаз
– ретинобластома – 80%
– отосклероз – 40%
– колотокома – 10%
– Пенетрантностъ отражает частоту фенотипического проявления имеющейся в генотипе информации. Она соответствует проценту особей, у которых доминантный аллель гена проявился в признак, по отношению ко всем носителям этого аллеля. Неполная пенетрантность доминантного аллеля гена может быть обусловлена системой генотипа, в которой функционирует данный аллель и которая является своеобразной средой для него. Взаимодействие неаллельных генов в процессе формирования признака может привести при определенном сочетании их аллелей к непроявлению доминантного аллеля одного из них.
Хорея Гентингтона проявляется в непроизвольном подергивании головы. Конечностей, постепенно прогрессирует и приводит к смерти. Может проявиться в раннем постэмбриональном периоде, в зрелом возрасте или не проявиться вообще. И экспрессивность, и пенетрантность поддерживаются естественным отбором, т.е. гены, контролирующие патологические признаки могут иметь разную экспрессивность и пенетрантность: заболевают не все носители гена, а у заболевших степень проявления будет различна. Проявление или неполное проявление признака, а так же его отсутствие зависит от среды и от модифицирующего действия других генов.
Ген может действовать плейотропно (множественно), т.е. опосредовано влиять на течение разных реакций и развитие многих признаков. Гены могут оказывать влияние на другие признаки на разных стадиях онтогенеза. Если ген включается в позднем онтогенезе, то оказывается незначительное действие. Если на ранних стадиях – изменения более значительны.
Фенилкетанурия. У больных есть мутация, которая выключает фермент – фенилаланин – гидролазу. Поэтому фенилаланин не превращается в тирозин. В результате в крови количество фенилаланина повышается. Если выявить эту патологию рано (до 1 месяца) и перевести ребенка на другое питание, развитие идет нормально, если позднее – понижен размер головного мозга, умственная отсталость, не развиваются нормально, отсутствует пигментация, умственные способности минимальны.
Плейотропность отражает интеграцию генов и признаков.
У человека есть патологический ген, приводящий к синдрому Фанкони (порок развития или отсутствие большого пальца, порок или отсутствие лучевой кости, недоразвитие почки, коричневые пигментные пятна, нехватка кровяных телец).
Есть ген, связанный с Х-хромосомой. Невосприимчивость к инфекциям и нехватка кровяных телец.
Доминантный ген, сцепленный с Х-хромосомой – пилонефрит, лабиринтная тугоухость.
Синдром Марфани – паучьи пальцы, вывих хрусталика глаза, пороки развития сердца.
Генокопия(греч. genos род, происхождение лат. copia множество) – термин в 1957 г. предложил
немецкий генетик Нахтсхейм (H.Nachtsheim).Обозначает сходные изменения одного и того же признака под влиянием разных неаллельных генов, которые иногда называют миметическими генами гетерогенной группы.
genocopies – генокопии.
Oдинаковые изменения фенотипа, обусловленные аллелями разных генов, а также имеющие место в результате различных генных взаимодействий или нарушений различных этапов одного биохимического процесса с прекращением синтеза конечного продукта, – например, у Drosophila melanogaster известен ряд мутаций неаллельных генов, обусловливающих фенотип “красные глаза” (нарушен синтез коричневого пигмента).
42.Изменчивость. Формы изменчивости: модификационная и генотипическая, их значение в онтогенезе и эволюции.
Изменчивость
Одним из признаков жизни является изменчивость. Любой живой организм отличается от других представителей вида
Изменчивость – свойство живых организмов существовать в разных формах. Групповая и индивидуальнаяизменчивость – классификация по эволюционному значению. Изменчивость, реализованная группой организмов, называется групповой, у одного организма или группы его клеток – индивидуальная.
По характеру изменения признаков и механизму:
–фенотипическая
– случайная
– модификационная
–генотипическая
– соматическая
– генеративная (мутационная, комбинативная)
а) генная
б) хромосомная
в) геномная
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера). Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Изменчивость бывает наследственная ( неопределенная, индивидуальная генотипическая) и ненаследственная ( определенная, групповая, модификационная). Наследственная изменчивость связана с изменением генотипа, ненаследственная- с изменением фенотипа под влиянием условий окружающей среды.
Значение мод.изм.:Адаптация- приспособление к данным условиям среды
Значение генотип. изм:Материал для естественного и искусственного отбора, распространение в популяции новых наследственных изменений.
43.Фенотипическая изменчивость и её виды. Модификации и их характеристики. Норма реакции признака. Фенокопии. Адаптивный характер модификаций.
По характеру изменения признаков и механизму:
–фенотипическая
– случайная
– модификационная
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера). Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Под влиянием внешних условий на генетически нормальный организм копируются признаки совсем другого генотипа. Проявление дальтонизма может произойти под влиянием питания, плохой психической конституции, повышенной раздражительности. У человека возникает заболевание витилиго (1% людей) – нарушение пигментации кожи. Генетический дефект есть у 30% болеющих, у остальных – профессиональное витилиго (воздействие на организм особых химических и отравляющих веществ). В Германии 15 лет назад рождались дети с фекомелией – укороченными ластовидными руками. Выяснилось. Что рождение таких детей происходило, если мать принимала Телидомид (успокоительное средство, показанное беременным). В результате нормальный немутантный генотип получал мутацию.
Фенокопии появляются в большинстве случаев при действии внешней среды на ранних стадиях эмбриогенеза, что приводит к врожденным заболеваниями порокам развития. Наличие фенокопий затрудняет диагностику заболеваний.
Норма реакции
Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет предел для каждого вида — например, усиленное кормление приведет к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и наследуется. Для разных изменений есть разные пределы нормы реакции. Например, сильно варьируют величина удоя, продуктивность злаков (количественные изменения), слабо — интенсивность окраски животных и т. д. (качественные изменения). В соответствии с этим норма реакции может быть широкой (количественные изменения — размеры листьев многих растений, размеры тела многих насекомых в зависимости от условий питания их личинок) и узкой (качественные изменения — окраска у куколок и имаго некоторых бабочек). Тем не менее, для некоторых количественных признаков характерна узкая норма реакции (жирность молока, число пальцев на ногах у морских свинок), а для некоторых качественных признаков — широкая (например, сезонные изменения окраски у многих видов животных северных широт).
Фенотипические изменения, возникающие на основе одного и того же генотипа в разных условиях его реализации, называют модификациями. Примером модификаций могут служить изменения содержания жира в молоке животных или массы тела в зависимости от их питания, изменения количества эритроцитов в крови, в зависимости от парциального давления кислорода в воздухе, изменения темпа роста растений при разной освещенности и содержании минеральных веществ в почве. Другим примером модификационной изменчивости являются различия, наблюдаемые у генетически идентичных монозиготных близнецов или потомков одного растения, полученных путем вегетативного размножения, но развивавшихся в разных условиях среды.
Модификации отдельного признака или свойства, формируемого данным генотипом, образуют непрерывный ряд. Частота встречаемости каждого варианта в таком вариационном ряду различна. Чаще обнаруживаются средние значения признака. Чем дальше признак отстоит от среднего значения, тем реже он наблюдается (рис. 6.1).
Так как фенотипическое проявление наследственной информации может модифицироваться условиями среды, в генотипе организма запрограммировано не конкретное значение отдельных его характеристик, а лишь возможность их формирования в определенных пределах, называемых нормой реакции. Таким образом, норма реакции представляет собой пределы модификационной изменчивости признака, допустимой при данном генотипе. Некоторые признаки характеризуются широкой нормой реакции. Как правило, это количественные признаки, контролируемые полигенами (масса тела, жирность молока, пигментация кожи), другие свойства характеризуются узкой нормой реакции и слабо или почти не модифицируются в разных условиях (цвет глаз, группа крови).
(Одним из признаков жизни является изменчивость. Любой живой организм отличается от других представителей вида. Изменчивость – свойство живых организмов существовать в разных формах. Фенотипическая изменчивость- ненаследственная, связана с изменением фенотипа под влиянием условий окружающей среды. Она может быть случайная и модификационная. Фенотипические изменения, возникающие на основе одного и того же генотипа в разных условиях его реализации- модификации. Пример: изменение содержания жира в молоке животных от их питания и т.д. Свойства:
1.возникают постепенно, имеют переходные формы
2. Модификации- изменения количественные, они образуют непрерывные ряды и группируются вокруг среднего значения.
3. Возникают направленно- под влиянием одного и того же фактора среды.
4. Модификации обратимы.
5. не передаются по наследству.
Фенотипические проявления наследственной информации может модифицироваться условиями среды, в генотипе организма запрограммировано не конкретное значение отдельных его признаков, а лишь возможность их формирования в определенных пределах- норма реакции. Норма реакции –это предел мод. изм.
Значение модификаций: позволяют адаптироваться к условиям внешней среды.
Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Под влиянием внешних условий на генетически нормальный организм копируются признаки совсем другого генотипа. Проявление дальтонизма может произойти под влиянием питания, плохой психической конституции, повышенной раздражительности. У человека возникает заболевание витилиго (1% людей) – нарушение пигментации кожи. Генетический дефект есть у 30% болеющих, у остальных – профессиональное витилиго (воздействие на организм особых химических и отравляющих веществ). В Германии 15 лет назад рождались дети с фекомелией – укороченными ластовидными руками. Выяснилось. Что рождение таких детей происходило, если мать принимала Телидомид (успокоительное средство, показанное беременным). В результате нормальный немутантный генотип получал мутацию. Фенокопии появляются в большинстве случаев при действии внешней среды на ранних стадиях эмбриогенеза, что приводит к врожденным заболеваниями порокам развития. Наличие фенокопий затрудняет диагностику заболеваний. )
44.Фенотип. Фенотип как результат реализации наследственной информации (генотипа) в определенных условиях среды. Значение средовых и генотипических факторов в формировании патологически измененного фенотипа человека.
Генотип – совокупность всех генов организма (генетическая конституция).
Фенотип – все признаки организма, формирующиеся в результате взаимодействия генотипа и среды. (Иогансен – 1803год) свойства любого организма зависят от генотипа и от среды, поэтому формирование организма – результат взаимодействия генетических факторов и факторов внешней среды.
Долгое время считалось, что в зиготе находятся разные хромосомы для разных клеток, однако теперь известно, что в зиготе имеется та же генетическая информация, что и во всех клетках данного организма. В специализированных клетках работают гены, характерные для функций данных клеток, а все остальные – до 95% – заблокированы. Каждая эмбриональная клетка потенциально может стать любой клеткой организма, т.е. специализироваться в любую сторону – полипотентные клетки. Каждая клетка организма способна дифференцироваться только по одному пути. Направление специализации определяется внешней средой (химическим окружением хромосом – цитоплазмой). На самых ранних этапах эмбриогенеза, генотип уже взаимодействует со средой. Взаимодействие удобно просматривать на примере глобиновых генов. До и после рождения эти гены работают неодинаково. В раннем эмбриогенезе включается ген, отвечающий за альфа-цепь гемоглобина (он активен на протяжении всей жизни), а ген, отвечающий за синтез бета-цепи, неактивен. Зато есть ген, отвечающий за синтез гамма-цепи. После рождения ген бета-цепи начинает работать, а гамма – блокируется. Эти изменения связаны с особенностями дыхания. Фетальный гемоглобин легко доносит воздух до зародыша.
Фенотипическое проявление генотипа в зависимости Ио среды изменяется в пределах нормы реакции. От родителей потомки получают специфические типы химических реакций на разные условия среды. Совокупность всех химических реакций определят метаболизм – обмен веществ. Интенсивность обмена веществ варьирует в широких пределах. У каждого человека свои особенности обмена веществ, которые передается от поколения к поколению, и подчиняются законам Менделя. Различия в обмене веществ реализуются в конкретных условиях среды на уровне синтеза белка.
Дифференцированная реакция растений примулы в разных условиях окружающей среды. При обычной температуре 20-25 градусов и нормальном давлении – красные цветы, при повышенной температуре или давлении – белые цветы. Семена обладают теми же свойствами.
Муха – дрозофила имеет ген, формирующий замыкание крыльев на спину. Если мух с мутантным генов выводить при температуре22-25 градусов, крылья загнуты. При более низкой температуре – нормальные крылья и лишь у некоторых – загнуты. Ген обуславливает синтез термочувствительного белка. Поэтому, обсыхая после выхода из куколки, при повышенной температуре происходит деформация крыльев.
Никакие признаки не наследуются. Признаки развиваются на основе взаимодействия генотипа и среды. Наследуется только генотип, т.е. комплекс генов, который определяет норму биологической реакции организма, изменяющую проявление и выраженность признаков в разных условиях среды. Таким образом, организм реагирует на свойства внешней среды. Иногда один и тот же ген в зависимости от генотипа и от условий внешней среды по-разному проявляет признак или меняет полноту выраженности.
45.Комбинативная изменчивость, её механизмы. Значение комбинативной изменчивости в обеспечении генотипического разнообразия людей.
Комбинативная изменчивость – результат независимого расхождения хромосом в процессе мейоза, оплодотворения, кроссинговера с рекомбинацией генов. При комбинативной изменчивости происходит перекомбинация генов, возникает новый индивидуальный набор хромосом, а значит, новый генотип и фенотип. Источником комбинативной изменчивостиявляются следующие процессы, происходящие в ходе мейоза и в результате оплодотворения: рекомбинация генов в результате кроссинговера, рекомбинация хромосом в ходе мейоза, комбинации хромосом, возникающие в результате слияния гамет при оплодотворении. К. И., проявляющаяся в генотипическом разнообразии особей, повышает выживаемость вида в изменяющихся условиях. Для комбинативной изменчивости в системе людей большое значение имеет система браков. Самая простая – случайный подбор пар (панмиксия). Строго панмиксных популяций не существует, т.к. существуют ограничения: социальные, религиозные, индивидуальные, экономические и другие.
46.Генные болезни человека, механизмы их возникновения и проявления. Примеры. С 258-261
Генные болезни – это большая группа заболеваний, возникающих в результате повреждения ДНК на уровне гена.
В зависимости от функциональной значимости первичных продуктов соответствующих генов генные болезни подразделяют на наследственные нарушения ферментных систем (энзимопатии), дефекты белков крови (гемоглобинопатии), дефекты структурных белков (коллагеновые болезни) и генные болезни с невыясненным первичным биохимическим дефектом.
Энзимопатии. В основе энзимопатии лежат либо изменения активности фермента, либо снижение интенсивности его синтеза. У гетерозигот-носителей мутантного гена присутствие нормального аллеля обеспечивает сохранение около 50% активности фермента по сравнению с нормальным состоянием. Поэтому наследственные дефекты ферментов клинически проявляются у гомозигот, а у гетерозигот недостаточная активность фермента выявляется специальными исследованиями.
В зависимости от характера нарушения обмена веществ в клетках среди энзимопатий различают следующие формы.
1. Наследственные дефекты обмена углеводов (галактоземия — нарушение метаболизма молочного сахара — лактозы; мукополисаха-ридозы — нарушение расщепления полисахаридов).
2. Наследственные дефекты обмена липидов и липопротеинов (сфинголипидозы — нарушение расщепления структурных липидов; нарушения обмена липидов плазмы крови, сопровождающиеся увеличением или снижением в крови холестерина, лецитина).
3. Наследственные дефекты обмена аминокислот (фенилкетонурия —нарушение обмена фенилаланина (см. разд. 4.1); тирозиноз— нарушение обмена тирозина; альбинизм — нарушение синтеза пигмента меланина из тирозина и др.).
4. Наследственные дефекты обмена витаминов (гомоцистинурия — развивается как результат генетического, дефекта кофермента витаминов В6 и B12, наследуется по аутосомно-рецессивному типу).
5. Наследственные дефекты обмена пуриновых и пиримидиновых азотистых оснований (синдром Леша — Найяна, связанный с недостаточностью фермента, который катализирует превращение свободных пуриновых оснований в нуклеотиды, наследуется по Х-сцепленному рецессивному типу).
6. Наследственные дефекты биосинтеза гормонов (адреногенитальный синдром, связанный с мутациями генов, которые контролируют синтез андрогенов; тестикулярная феминизация, при которой не образуются рецепторы андрогенов).
7. Наследственные дефекты ферментов эритроцитов (некоторые гемолитические несфероцитарные анемии, характеризующиеся нормальной структурой гемоглобина, но нарушением ферментной системы, участвующей в анаэробном (бескислородном) расщеплении глюкозы. Наследуются как по аутосомно-рецессивному, так и по Х-сцепленному рецессивному типу).
Гемоглобинопатии. Это группа наследственных заболеваний, вызываемых первичным дефектом пептидных цепей гемоглобина и связанным с этим нарушением его свойств и функций. К ним относят метгемоглобинемии, эритроцитозы, серповидно-клеточную анемию, талассемии (см. § 4.1).
Коллагеновые болезни. В основе возникновения этих заболеваний лежат генетические дефекты биосинтеза и распада коллагена — важнейшего структурного компонента соединительной ткани. К этой группе относят болезнь Эллерса — Данлоса, характеризующуюся большим генетическим полиморфизмом и наследующуюся как по аутосомно-доминантному, так и по аутосомно-рецессивному типу, болезнь Марфана, наследующуюся по аутосомно-доминантному типу, и ряд других заболеваний.
Наследственные болезни с невыясненным первичным биохимическим дефектом. К этой группе принадлежит подавляющее большинство моногенных наследственных болезней. Наиболее распространенными являются следующие.
1. Муковисцидозы — встречаются с частотой 1:2500 новорожденных; наследуются по аутосомно-рецессивному типу. В основе патогенеза заболевания —наследственное поражение экзокринных желез и железистых клеток организма, выделение ими густого, измененного по составу секрета и связанные с этим последствия.
2. Ахондроплазия — заболевание, в 80—95% случаев обусловленное вновь возникшей мутацией; наследуется по аутосомно-доминантному типу; встречается с частотой приблизительно 1:100 000. Это заболевание костной системы, при котором наблюдаются аномалии развития хрящевой ткани преимущественно в эпифизах трубчатых костей и костях основания черепа (рис. 6.23).
3. Мышечные дистрофии (миопатии) —заболевания, связанные с поражением поперечно-полосатых и гладких мышц. Различные формы характеризуются разным типом наследования. Например, прогрессирующая псевдогипертрофическая миопатия Дюшена наследуется по Х-сцепленному рецессивному типу и проявляется преимущественно у мальчиков в начале первого десятилетия жизни. Известна мышечная псевдогипертрофическая дистрофия, наследующаяся по аутосомно-рецессивному типу, которая начинает развиваться во второй половине первого десятилетия жизни и встречается с одинаковой частотой у обоих полов. Мышечная дистрофия плечевого и тазового пояса: наследуется по аутосомно-доминантному типу и т.д.
Генетическое многообразие генных болезней. Изучение наследственных заболеваний у человека свидетельствует о том, что нередко сходное фенотипическое проявление болезни бывает обусловлено несколькими различными мутациями. Это явление впервые было описано в 30-х гг. С. Н. Давиденковым и названо генетической гетерогенностью наследственных заболеваний. Генетическая гетерогенность наследственных болезней может быть обусловлена мутациями разных генов, кодирующих ферменты одного метаболического пути, а также мутациями одного и того же гена, приводящими к появлению разных его аллелей.
Среди рассмотренных выше наследственных болезней особенно высокой степенью генетического полиморфизма отличаются мукопо-лисахаридозы, генетическая разнородность которых объясняется множественными мутациями в 11—12 генах, связанных общей функцией расщепления полисахаридов. Большой генетической гетерогенностью характеризуется врожденная аутосомно-рецессивная форма глухоты, при которой различают не менее 35 генетически различных вариантов с фенотипически сходным проявлением.
Большие перспективы в расшифровке наследственной гетерогенности генных болезней открываются в связи с применением молекулярно-генетических методов их прямого анализа с помощью ДНК-зондов.
Есть моногенные и полигенные болезни.
Моногенные болезни наследственного предрасположения – наследственные заболевания, проявляющиеся из-за мутации одного гена или проявляющиеся при действии определенного фактора среды (аутосомно-рецессивные или сцепленные с Х-хромосомой).
Проявляются при воздействии факторов:
– физических;
– химических;
– пищевых;
– загрязнения среды.
Парамиотомия – в сырую погоду происходят тонические спазмы мышц при холоде, под влиянием тепла – проходят. Болезнь связана с термочувствительным белком. Реакция проявляется в младенчестве и не изменяется на протяжении жизни человека.
Пигментная ксеродерма – веснушчатая кожа особого типа. Проявляется в 4-6 лет. Дети не переносят УФ-свет возникают злокачественные опухоли, такие дети умирают от метастаз еще до 15 лет. Не переносят также и гамма-лучей.
Синдром Блюма. Пигментная «бабочка» на лице, маленький рост, удлиненная голова. Евреи, поляки, беларусы, австрийцы. Погибают до 18 лет. Не переносят УФ-облучения, гамма-лучей.
Альфа-1 антитрипсин при загрязнении воздуха, табачном дыме проявляется острой закупоркой бронхов или циррозом печени.
У европеоидов люди, не переносящие молоко, составляют 10-20%, в Африке – 70-80%.
Влияние лекарственных средств: сульфаниламидные препараты провоцируют заболевания крови.
Есть полигенные болезни наследственного происхождения – такие болезни, которые возникают при действии многих факторов (мультифакториальные) и в результате взаимодействия многих генов. Установить диагноз в таком случае очень сложно, т.к. действует много факторов, и появляется новое качество при взаимодействии факторов.
Хромосомные болезни человека, механизмы их возникновения и проявления. Примеры.
Хромосомные болезни, наследственные заболевания, обусловленные изменением числа или структуры хромосом.
Эта группа заболеваний обусловлена изменением структуры отдельных хромосом или их количества в кариотипе. Как правило, при таких мутациях наблюдается дисбаланс наследственного материала, который и ведет к нарушению развития организма. У человека описаны геномные мутации по типу полиплоидии, которые редко наблюдаются у живорожденных, а в основном обнаруживаются у абортированных эмбрионов и плодов и у мертворожденных. Основную часть хромосомных болезней составляют анэуплоидии, причем моносомии по аутосомам у живорожденных встречаются крайне редко. Большинство из них касаются 21-й и 22-й хромосом и чаще обнаруживаются у мозаиков, имеющих одновременно клетки с нормальным и мутантным кариотипом. Достаточно редко обнаруживается моносомия и по Х-хромосоме (синдром Шерешевского — Тернера).
В отличие от моносомии трисомии описаны по большому числу аутосом: 8, 9, 13, 14, 18, 21, 22-й и Х-хромосоме, которая может присутствовать в кариотипе в 4—5 экземплярах, что вполне совместимо с жизнью.
Структурные перестройки хромосом также, как правило, сопровождаются дисбалансом генетического материала (делеции, дупликации). Степень снижения жизнеспособности при хромосомных аберрациях зависит от количества недостающего или избыточного наследственного материала и от вида измененной хромосомы.
К настоящему времени описано около 100 клинико-цитогенетических синдромов, в основе которых лежат различные хромосомные аномалии.
Хромосомные изменения, приводящие к порокам развития, чаще всего привносятся в зиготу с гаметой одного из родителей при оплодотворении. При этом все клетки нового организма будут содержать аномальный хромосомный набор и для диагностики такого заболевания достаточно проанализировать кариотип клеток какой-нибудь ткани.
Если хромосомные нарушения возникают в одном из бластомеров во время первых делений зиготы, образующейся из нормальных гамет, то развивается мозаичный организм, большая или меньшая часть клеток которого несет нормальный хромосомный набор. Диагностика мозаичных форм хромосомных болезней отличается большей трудоемкостью и требует изучения кариотипа большого числа клеток из разных тканей.
Для определения вероятности появления хромосомной болезни в потомстве в семьях, уже имеющих больных детей, важно установить, является ли это хромосомное нарушение заново возникшим или оно унаследовано от предыдущего поколения. Чаще родители человека с хромосомным заболеванием имеют нормальный кариотип, а появление больного потомства является результатом мутации, возникшей в одной из гамет. В этом случае возможность повторного хромосомного нарушения у детей в данной семье маловероятна и не превосходит таковой в целом для популяции. Вместе с тем описано немало семей, в которых наблюдается предрасположение, например, к нерасхождению хромосом.
В случае наследуемых хромосомных болезней в соматических клетках родителей обнаруживаются хромосомные или геномные мутации, которые могут передаваться их зрелым половым клеткам в ходе гаметогенеза. Передают потомству хромосомные нарушения обычно фенотипически нормальные родители, являющиеся носителями сбалансированных хромосомных перестроек — реципрокных транслокаций, робертсоновских транслокаций или перицентрических инверсий. У носителей такого рода хромосомных перестроек с определенной вероятностью образуются нормальные гаметы, а также гаметы, несущие сбалансированную перестройку, и половые клетки с нарушенным балансом генов в геноме (рис. 6.22).

Рис. 6.22. Вероятность образования нормальных и аномальных гамет у носителей сбалансированной хромосомной перестройки
Показана робертсоновская транслокация 21-й хромосомы (окрашена) на одну из акродентрических хромосом (не окрашена);
I — кариотип носителя сбалансированной хромосомной перестройки, II — варианты (а, б) расположения бивалентов в экваториальной плоскости веретена деления (метафаза I), III — результат 1-го редукционного деления мейоза; IV — результат 2-го эквационного деления мейоза; V — гаметы со сбалансированной хромосомной перестройкой; VI — нормальные гаметы; VII — гаметы, не имеющие 21-й хромосомы; VIII — гаметы, содержащие две 21-е хромосомы (VII и VIII — гаметы с несбалансированным геномом)
Возможность наследования хромосомных аномалий делает необходимым анализ кариотипа родителей, уже имеющих больных детей, и пренатальную диагностику развивающегося внутриутробно плода для исключения вероятности повторного рождения ребенка с хромосомной болезнью.
Фенотипическое проявление различных хромосомных и геномных мутаций характеризуется ранним и множественным поражением различных систем органов. Типичными являются задержка общего физического и умственного развития, отклонения в строении скелета, в частности мозгового и лицевого черепа, пороки развития сердечно-сосудистой, мочеполовой, нервной систем, нарушения в биохимическом, гормональном и иммунологическом статусе организма. Хромосомные болезни, как правило, характеризуются сочетанием многих врожденных пороков. Для них также характерны многообразие и вариабельность фенотипических проявлений. Наиболее специфические проявления хромосомных заболеваний связаны с дисбалансом по относительно небольшому фрагменту хромосомы. Так, фенотипическое проявление синдрома Дауна наблюдается в случае трисомии всего лишь по небольшому сегменту длинного плеча 21-й хромосомы. Картина синдрома «кошачьего крика» развивается при утрате участка короткого плеча 5-й хромосомы. Дисбаланс по значительному объему хромосомного материала делает фенотипическую картину менее специфической.
Специфичность проявления хромосомного заболевания определяется изменением содержания определенных структурных генов, кодирующих синтез специфических белков. Так, при болезни Дауна обнаружено повышение в 1,5 раза активности фермента супероксид-дисмутазы I, ген которого располагается в 21-й хромосоме и представлен у больных в трехкратной дозе. Эффект «дозы гена» обнаружен более чем для 30 генов, локализованных в разных хромосомах человека.
Полуспецифические симптомы проявления хромосомных болезней связаны в значительной мере с дисбалансом генов, представленных многими копиями, которые контролируют ключевые процессы в жизнедеятельности клеток и кодируют, к примеру, структуру рРНК, тРНК, гистонов, рибосомальных белков, актина, тубулина.
Неспецифические проявления при хромосомных болезнях связывают с изменением содержания гетерохроматина в клетках, который оказывает влияние на нормальное течение клеточного деления и роста, формирование в онтогенезе количественных признаков, определяемых полигенами.
Для медицинской практики в 1971 году был проведен симпозиум по медицинской генетике в Париже. Была принята международная Парижская классификация для обозначения кариотипа человека. 46,хх; 46,ху – кариотип нормального человека.
Во время мейоза возможно появление аномальных половых клеток.
47,хху – синдром Клайнфельтера.
Мужчина, частота встречаемости 1 из 1000 новорожденных мальчиков.
§
синдром Дауна — трисомия по 21 хромосоме, к признакам относятся: слабоумие, задержка роста, характерная внешность, изменения дерматоглифики;
синдром Патау — трисомия по 13 хромосоме, характеризуется множественными пороками развития, идиотией, часто — полидактилия, нарушения строения половых органов, глухота; практически все больные не доживают до одного года;
синдром Эдвардса — трисомия по 18 хромосоме.
Болезни, связанные с нарушением числа половых хромосом
– синдром Шерешевского-Тернера — отсутствие одной Х-хромосомы у женщин (45 ХО) вследствие нарушения расхождения половых хромосом; к признакам относится низкорослость, половой инфантилизм и бесплодие, различные соматические нарушения (микрогнатия, короткая шея и др.);
– полисомия по Х-хромосоме — включает трисомию (кариотии 47, XXX), тетрасомию (48, ХХХХ), пентасомию (49, ХХХХХ), отмечается незначительное снижение интеллекта, повышенная вероятность развития психозов и шизофрении с неблагоприятным типом течения;
– синдром Кляйнфельтера — полисомия по X- и Y-хромосомам у мальчиков (47, XXY; 47, XYY, 48, XXYY и др.), признаки: евнухоидный тип сложения, гинекомастия, слабый рост волос на лице, в подмышечных впадинах и на лобке, половой инфантилизм, бесплодие; умственное развитие отстает, однако иногда интеллект нормальный.
Болезни, причиной которых является полиплоидия
· триплоидии, тетраплоидии и т. д.; причина — нарушение процесса мейоза вследствие мутации, в результате чего дочерняя половая клетка получает вместо гаплоидного (23) диплоидный (46) набор хромосом, то есть 69 хромосом (у мужчин кариотип 69, XYY, у женщин — 69, XXX); почти всегда летальны до рождения.
Нарушения структуры хромосом
· Транслокации — обменные перестройки между негомологичными хромосомами.
· Делеции — потери участка хромосомы. Например, синдром «кошачьего крика» связан с делецией короткого плеча 5-ой хромосомы. Признаком его служит необычный плач детей, напоминающий мяуканье или крик кошки. Это связано с патологией гортани или голосовых связок. Наиболее типичным, помимо «кошачьего крика», является умственное и физическое недоразвитие, микроцефалия (аномально уменьшенная голова).
· Инверсии — повороты участка хромосомы на 180 градусов.
· Дупликации — удвоения участка хромосомы.
· Изохромосомия — хромосомы с повторяющимся генетическим материалом в обоих плечах.
· Возникновение кольцевых хромосом — соединение двух концевых делеций в обоих плечах хромосомы.
В настоящее время у человека известно более 700 заболеваний, вызванных изменением числа или структуры хромосом. Около 25 % приходится на аутосомные трисомии, 46 % — на патологию половых хромосом. Структурные перестройки составляют 10,4 %. Среди хромосомных перестроек наиболее часто встречаются транслокации и делеции.
Геномные болезни человека, механизмы их возникновения и проявления. Примеры.
47,хху – синдром Клайнфельтера.
Мужчина, частота встречаемости 1 из 1000 новорожденных мальчиков.
Высокий рост, более длинные ноги, евнуховидное телосложение, недоразвитие половых органов, гинекомастия, у половины умственная отсталость (трудности в обучении чтению и письму), могут заканчивать нормальные школы, хотя им может быть очень трудно. Вспыльчивы, импульсивны, легко попадают од влияние более сильных личностей, преступления и проступки. Жизнеспособность снижена. Среди «туповатых» преступников приблизительно 2%.
Хуу – синдром двойного игрек (трисомия)
1 на 700 новорожденных. Впервые в 1977году были исследованы.
Высокие мужчины, агрессивное поведение, интеллект снижен или находится на нижней границе нормы. Характерные преступления – поджоги, воровство, детоубийство без мотивации. В больницах закрытого типа, в колонии – 5% таких людей. Поведение детерминировано лишней хромосомой.
Ххх – синдром Сверхженщины.
1на 1000 новорожденных девочек.
Внешне не проявляется, легкое слабоумие. Считают, что около 1% девушек и женщин с легким слабоумием. Могут беременеть и рождают нормальных детей (во время мейоза происходит самокоррекция).
45,у0 – нежизнеспособны – аборт.
Х0 синдром Шеришевкого-Тернера
частота встречаемости 1:2000 девочек. Летальность при моносомии очень высокая, каждый 13 выкидыш имеет такую природу. Фенотипические проявления – маленький рост, для многих характерна шейная складка. Локтевой изгиб под углом, укорочены 4 и 5 пальцы, антимонголоидные глаза, абстрактное мышление отсутствует, упорные, трудолюбивые, способны заканчивать школы, ВУЗы. Любовь к опеканию маленьких детей. Отсутствует критическое восприятие своих дефектов. Низкий рост девочки – непременное условие для проведения кариотипирования. Окружность головы больше нормы, груди широко расставлены.
49,ххххх – нарушения те же, Но встречаемость ниже 49,хххху – то же.
Аутосом меньше 44 не бывает, но больше – возможно.
49.Болезни человека с наследственной предрасположенностью, механизмы их возникновения и проявления. Примеры. С 262-263.
50.Значение генетики для медицины. Человек как специфический объект генетического анализа. Методы изучения наследственности человека: близнецовый, генеалогический., цитогенетический, биохимический, популяционно-статистический, генетики соматических клеток, методы изучения ДНК.
Генетика человека – основа медицинской генетики. Человек – удобный генетический объект. У человека лучше, чем у других видов изучены биохимические, иммунологические, физиологические и другие реакции, а эти признаки детерминированы генами. Генетика человека – наука о наследственно обусловленных различиях людей и о нарушениях генетического материала. Существует специальность – клиническая генетика Клиническая генетика – дисциплина, которая применяет законы классической генетики к проблемам клинической медицины, изучает этиологию и патогенез наследственных заболеваний, разрабатывает методы лечения наследственных заболеваний.
Основные закономерности наследственности были открыты благодаря применению гибридологического метода (Мендель). Для человека невозможен, так как:
1) У человека не может быть произведено искусственного скрещивания
2) Низкая плодовитость делает невозможным применение статистического подхода при оценке немногочисленного потомства одой пары родителей
3) Редкая смена поколений при значительной продолжительности жизни дает возможность одному исследователю наблюдать не более 3-4 поколений.
Существуют разные методы генетики человека.
1. Генеалогический метод.
Анализ многих заболеваний приводит к выводу об их наследовании в семье. Семейный характер заболеваний можно выявить при анализе родословных метод анализа или изучения передачи признака среди родственников больного в ряду поколений. Родословную составляют по отношению к одному определенному признаку, наследование которого необходимо проследить.
Сущность генеалогического метода.
-установление наследственной этиологии заболевания
– установление типа наследования
– установление локализации гена в хромосоме
– дифференциация болезней генетической этиологии с близкими к ним ненаследственми формами
– дифференциация наследственных заболеваний, несходных по гены наследования.
2. Близнецовый метод.
Рождение близнецов – довольно редкое явление.
Монозиготные близнецы – редкое явление, не зависит от возраста, расы, географического положения и условий жизни. Имеют большое значение для анализа т.к. очень похожи. Близнецовый метод позволяет установить, в какой мере признак зависит от генов и в какой – от внешней среды (т.к. организм взаимодействует с окружающей средой). Используются понятия: конкордантность и дискордантность. Близнецовые методы часто используются для исследования туберкулеза, сахарного диабета, шизофрении, эпилепсии и других болезней и отклонений.
Биохимические методы.
В мире проводят более 700 исследований для клинических нужд, 300 для наследственных болезней. Болезни, которые имеют выявленные биохимические дефекты.
– болезни обмена аминокислот
– болезни обмена углеводов
– болезни обмена липидов
– болезни стероидного обмена
– болезни обмена пуриновых и пиримидиновых оснований и др.
Энзимопатии у 10000 ферментов.
Биохимические методы используются для определения генотипа носителей, установления генотипа родителей по данному признаку.
§
– равная вероятность встречаемости данного признака как у мужчин, так и у женщин. Это обусловлено одинаковой двойной дозой генов, расположенных в аутосомах у всех представителей вида и получаемых от обоих родителей, и зависимостью развивающегося признака от характера взаимодействия аллельных генов.
При доминировании признака в потомстве родительской пары, где хотя бы один родитель является его носителем, он проявляется с большей или меньшей вероятностью в зависимости от генетической конституции родителей (рис. 6.25).

Если анализируется признак, не влияющий на жизнеспособность организма, то носители доминантного признака могут быть как гомо-, так и гетерозиготами. В случае доминантного наследования какого-то патологического признака (заболевания) гомозиготы, как правило, нежизнеспособны, а носители этого признака — гетерозиготы.
Таким образом, при аутосомно-доминантном наследовании признак может встречаться в равной мере у мужчин и у женщин и прослеживается при достаточном по численности потомстве в каждом поколении по вертикали. Анализируя родословные, необходимо помнить о возможности неполного пенетрирования доминантного аллеля, обусловленной взаимодействием генов или факторами среды. Показатель пенетрантности может быть вычислен как отношение фактического числа носителей признака к числу ожидаемых носителей этого признака в данной семье. Необходимо также помнить, что некоторые заболевания проявляются не сразу с момента рождения ребенка. Многие болезни, наследуемые по доминантному типу, развиваются лишь в определенном возрасте. Так, хорея Гентингтона клинически проявляется к 35—40 годам, поздно проявляется и поликистоз почек. Поэтому при прогнозировании подобных заболеваний в расчет не принимаются братья и сестры, не достигшие критического возраста.
Родословные при аутосомно-рецессивном наследовании. Рецессивные признаки проявляются фенотипически лишь у гомозигот по рецессивным аллелям. Эти признаки, как правило, обнаруживаются у потомков фенотипически нормальных родителей — носителей рецессивных аллелей. Вероятность появления рецессивного потомства в этом случае равна 25%. Если один из родителей имеет рецессивный признак, то вероятность проявления его в потомстве будет зависеть от генотипа другого родителя. У рецессивных родителей все потомство унаследует соответствующий рецессивный признак.
Для родословных при аутосомно-рецессивном типе наследования характерно, что признак проявляется далеко не в каждом поколении. Чаще всего рецессивное потомство появляется у родителей с доминантным признаком, причем вероятность появления такого потомства возрастает в близкородственных браках, где оба родителя могут являться носителями одного и того же рецессивного аллеля, полученного от общего предка.
Родословные при доминантном Х-сцепленном наследовании признака. Гены, расположенные в Х-хромосоме и не имеющие аллелей в Y-хромосоме, представлены в генотипах мужчин и женщин в разных дозах. Женщина получает две свои Х-хромосомы и соответствующие гены как от отца, так и от матери, а мужчина наследует свою единственную Х-хромосому только от матери. Развитие соответствующего признака у мужчин определяется единственным аллелем, присутствующим в его генотипе, а у женщин он является результатом взаимодействия двух аллельных генов. В связи с этим признаки, наследуемые по Х-сцепленному типу, встречаются в популяции с разной вероятностью у мужского и женского пола.
При доминантном Х-сцепленном наследовании признак чаще встречается у женщин в связи с большей возможностью получения ими соответствующего аллеля либо от отца, либо от матери. Мужчины могут наследовать этот признак только от матери. Женщины с доминантным признаком передают его в равной степени дочерям и сыновьям, а мужчины — только дочерям. Сыновья никогда не наследуют от отцов доминантного Х-сцепленного признака.
Примером такого типа наследования служит описанная в 1925 г. родословная с фолликулярным кератозом —кожным заболеванием, сопровождающимся потерей ресниц, бровей, волос на голове (рис. 6.30). Характерным является более тяжелое течение заболевания у гемизиготных мужчин, чем у женщин, которые чаще всего являются гетерозиготами.
При некоторых заболеваниях наблюдается гибель мужчин-гемизигот на ранних стадиях онтогенеза. Тогда в родословных среди пораженных должны быть только женщины, в потомстве которых отношение пораженных дочерей, здоровых дочерей и здоровых сыновей равно 1:1:1. Мужские доминантные гемизиготы, не погибающие на очень ранних стадиях развития, обнаруживаются в самопроизвольных абортах или среди мертворожденных. Такими особенностями наследования у человека характеризуется пигментный дерматоз.
Родословные при рецессивном Х-сцепленном наследовании признаков. Характерной особенностью родословных при данном типе наследования является преимущественное проявление признака у гемизиготных мужчин, которые наследуют его от матерей с доминантным фенотипом, являющихся носительницами рецессивного аллеля. Как правило, признак наследуется мужчинами через поколение от деда по материнской линии к внуку. У женщин он проявляется лишь в гомозиготном состоянии, вероятность чего возрастает при близкородственных браках.
Наиболее известным примером рецессивного Х-сцепленного наследования является гемофилия.
Другим примером наследования по данному типу является дальтонизм — определенная форма нарушения цветоощущения.
Родословные при Y-сцепленном наследовании. Наличие Y-хромосомы только у представителей мужского пола объясняет особенности Y-сцепленного, или голандриче-ского, наследования признака, который обнаруживается лишь у мужчин и передается по мужской линии из поколения в поколение от отца к сыну.
Одним из признаков, Y-сцепленное наследование которого у человека все еще обсуждается, является гипертрихоз ушной раковины, или наличие волос на внешнем крае ушной раковины. Предполагают, что в коротком плече Y-хромосомы кроме этого гена находятся гены, определяющие мужской пол. В 1955 г. у мыши описан определяемый Y-хромосомой трансплантационный антиген, названный HY. Возможно, он является одним из факторов половой дифференцировки мужских гонад, клетки которых имеют рецепторы, связывающие этот антиген. Связанный с рецептором антиген активизирует развитие гонады по мужскому типу (см. разд. 3.6.5.2; 6.1.2). Этот антиген в процессе эволюции остался почти неизменным и встречается в организме многих видов животных, в том числе и человека. Таким образом, наследование способности к развитию гонад по мужскому типу определяется голандрическим геном, расположенным в Y-хромосоме (рис. 6.32).
Сущность генеалогического метода.
-установление наследственной этиологии заболевания
– установление типа наследования
– установление локализации гена в хромосоме
– дифференциация болезней генетической этиологии с близкими к ним ненаследственми формами
– дифференциация наследственных заболеваний, несходных по гены наследования.
53.Близнецовый метод изучения генетики человека, возможности метода. Определение соотносительной роли наследственности и среды в развитии признаков и патологических состояний человека.
Рождение близнецов – довольно редкое явление. В европейских странах 1-1,2% всех родов – близнецы, в Восточной Европе РФ 1,5%, в Нигерии 4%, в Японии 0,7%
у европеоидов – 0,7%
у негроидов – 1,1%
у монголоидов – 0,2%
Эти различия касаются дизиготных близнецов. Зависит от возраста матери (чем старше, тем больше вероятность рождения близнецов), искусственное оплодотворение.
Монозиготные близнецы – редкое явление, не зависит от возраста, расы, географического положения и условий жизни. Имеют большое значение для анализа т.к. очень похожи. Близнецовый метод позволяет установить, в какой мере признак зависит от генов и в какой – от внешней среды (т.к. организм взаимодействует с окружающей средой). Используются понятия: конкордантность и дискордантность. Близнецовые методы часто используются для исследования туберкулеза, сахарного диабета, шизофрении, эпилепсии и других болезней и отклонений.
Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов. Он предложен в 1875 г. Гальтоном первоначально для оценки роли наследственности и среды в развитии психических свойств человека. В настоящее время этот метод широко применяют в изучении наследственности и изменчивости у человека для определения соотносительной роли наследственности и среды в формировании различных признаков, как нормальных, так и патологических. Он позволяет выявить наследственный характер признака, определить пенетрантность аллеля, оценить эффективность действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания).
Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны, так как имеют 100% общих генов. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордантных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбрионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам между близнецами наблюдается дискордантность, т.е. различия. Напротив, сохранение сходства между близнецами, несмотря на различия условий их существования, свидетельствует о наследственной обусловленности признака.
Сопоставление парной конкордантности по данному признаку у генетически идентичных монозиготных и дизиготных близнецов, которые имеют в среднем около 50% общих генов, дает возможность более объективно судить о роли генотипа в формировании признака. Высокая конкордантность в парах монозиготных близнецов и существенно более низкая конкордантность в парах дизиготных близнецов свидетельствуют о значении наследственных различий в этих парах для определения признака. Сходство показателя конкордантности у моно- и дизиготных близнецов свидетельствует о незначительной роли генетических различий и определяющей роли среды в формировании признака или развития заболевания. Достоверно различающиеся, но достаточно низкие показатели конкордантности в обеих группах близнецов дают возможность судить о наследственной предрасположенности к формированию признака, развивающегося под действием факторов среды.
Установление соотносительной роли наследственности и среды в развитии различных патологических состояний позволяет врачу правильно оценить ситуацию и проводить профилактические мероприятия при наследственной предрасположенности к заболеванию или осуществлять вспомогательную терапию при его наследственной обусловленности.
Трудности близнецового метода связаны, во-первых, с относительно низкой частотой рождения близнецов в популяции (1:86—1:88), что осложняет подбор достаточного количества пар с данным признаком; во-вторых, с идентификацией монозиготности близнецов, что имеет большое значение для получения достоверных выводов.
Для идентификации монозиготности близнецов применяют ряд методов. 1. Полисимптомный метод сравнения близнецов по многим морфологическим признакам (пигментации глаз, волос, кожи, форме волос и особенностям волосяного покрова на голове и теле, форме ушей, носа, губ, ногтей, тела, пальцевым узорам). 2. Методы, основанные на иммунологической идентичности близнецов по эритроцитарным антигенам (системы АВО, MN, резусу), по сывороточным белкам (γ-глобулину). 3. Наиболее достоверный критерий монозиготности предоставляет трансплантационный тест с применением перекрестной пересадки кожи близнецов.
Несмотря на трудоемкость близнецового метода и возможность ошибок при определении монозиготности близнецов, высокая объективность выводов делает его одним из широко применяемых методов генетических исследований у человека.
54.Цитогенетический метод изучения генетики человека. Денверская и Парижская классификация хромосом. Возможности идентификации хромосом человека.
55.Медико-генетические аспекты брака. Близкородственные браки. Медико-генетическое консультирование
Существуют методы пренатальной диагностики. Как правило, пренатальный диагноз ставится в медико-генетических учреждениях. В России 7 центров федерального значения, 10 областных центров, 85 пунктов медико-генетического консультирования.
Первая медицинская консультация появилась в 20х годах в Москве благодаря невропатологу Девиденко. Почти все медико-генетические консультации связаны с нервными болезными. Первая медико-генетическая консультация в США возникла при Мачиганском университете в 1941 году. В Воронеже подобная консультация появилась в 1960 году под руководством невропатологов.
Медико-генетические консультации обслуживают примерно 1.5 миллионов человек каждая. Врач- генетик затрачивает на прием одного больного около 1,5 – 2 часов, при повторном обращении 30-40 минут.
Для подавляющего большинства наследственных болезней эффективных способов лечения не существует. Из этого следует, что в борьбе с наследственной патологией основная роль отводится профилактике рождения аномального потомства. Общий профилактический характер носят мероприятия, направленные на оздоровление окружающей среды, способствующие снижению ее мутагенного воздействия на наследственный материал человеческого организма. В последние десятилетия распространенным и эффективным способом профилактики наследственных болезней является медико-генетическое консультирование.
Медико-генетическое консультирование — это один из видов специализированной помощи населению, направленной в первую очередь на предупреждение появления в семье детей с наследственной патологией. С этой целью составляют прогноз рождения в данной семье ребенка с наследственной болезнью, родителям объясняют вероятность этого события и оказывают помощь в принятии решения. В случае большой вероятности рождения больного ребенка родителям рекомендуют либо воздержаться от деторождения, либо провести пренатальную диагностику, если она возможна при данном виде патологии.
Консультирование семей, обращающихся к врачу-генетику, включает три основных этапа. Как правило, за консультацией обращаются семьи, где уже имеется ребенок с наследственной патологией, или семьи, в которых имеются больные родственники. На первом этапе консультирования производится уточнение диагноза, что является необходимой предпосылкой любого консультирования. Уточнение диагноза в медико-генетической консультации проводят с помощью генетического анализа. Для этой цели используют генеалогический, цитогенетический, биохимический и другие требуемые методы исследований, которым подвергаются пробанд и его родственники. Точный клинический и генетический диагноз заболевания позволяет установить степень генетического риска и выбор эффективных методов пренатальной диагностики и профилактического лечения.
На втором этапе консультирования делают прогноз потомства. Генетический риск может быть определен либо путем теоретических расчетов, основанных на генетических закономерностях, либо с помощью эмпирических данных. Сущность генетического прогноза заключается в определении вероятности появления наследственной патологии в семье. Наиболее эффективным является проспективное консультирование, когда риск рождения больного ребенка определяют до наступления беременности или в ранние ее сроки. Такие консультации чаще проводят в случае кровного родства супругов, при отягощенной наследственности по линии мужа или жены, при воздействии вредных средовых факторов на супругов незадолго до наступления беременности. Ретроспективное консультирование проводят после рождения больного ребенка относительно здоровья будущих детей.
Определение прогноза потомства при разных формах наследственной патологии различно. При моногенных, менделирующих болезнях прогноз основывается на расчете вероятности появления потомства в соответствии с генетическими закономерностями. При этом, если известен тип наследования данного заболевания и по родословной удается установить генотип родителей, оценка риска сводится к анализу мевделевского расщепления. Если у пробанда установлена вновь возникшая мутация, то повторный риск рождения ребенка с такой же патологией незначителен.
Расчет риска при моногенном заболевании может осложниться при пониженной экспрессивности или неполной пенетрантности гена, позднем проявлении генетической аномалии, генетической гетерогенности заболевания и вообще в случае неточного диагноза.
При хромосомных болезнях определение риска повторного рождения потомства с хромосомными аномалиями зависит от того, нормальны ли кариотипы родителей, не обнаружено ли у них мозаицизма, не наблюдается ли семейной формы структурных аномалий хромосом. В случае отсутствия нарушений в кариотипе родителей вероятность повторного рождения второго ребенка с хромосомной аномалией оценивается по эмпирическим данным для каждого вида аномалии с учетом возраста родителей.
При мультифакториальных заболеваниях, т.е. заболеваниях с наследственным предрасположением, основой оценки риска являются эмпирические данные о популяционной и семейной частоте каждого из них.
Специфический генетический риск до 5% принято считать низким, до 10% —повышенным в легкой степени, до 20% —средним, выше 20% — высоким. Генетический риск средней степени расценивают как противопоказание к зачатию или показание к прерыванию уже имеющейся беременности. Возможность проведения пренатальной диагностики является определяющей для принятия положительного решения в отношении завершения беременности.
На третьем этапе консультирования врач-генетик в доступной форме объясняет семье степень генетического риска рождения наследственно аномального потомства, сущность пренатальной диагностики и помогает принять правильное решение в отношении деторождения. Однако окончательное решение этого вопроса остается за родителями.
Широкое использование медико-генетического консультирования, разработка способов пренатальной диагностики наследственных заболеваний позволяют существенно уменьшить вероятность появления потомства с наследственной патологией в отдельных семьях.
Чаще всего рецессивное потомство появляется у родителей с доминантным признаком, причем вероятность появления такого потомства возрастает в близкородственных браках, где оба родителя могут являться носителями одного и того же рецессивного аллеля, полученного от общего предка.
Инбридинг приводит к повышению постоянства фенотипических признаков в потомстве и, в конечном итоге, производится для получения линий генетически идентичных особей (инбредные линии), на которых удобно проводить биологические и медицинские эксперименты.
Как известно, организм получает каждый ген в двух экземплярах (аллелях) — от отца и от матери. Если эти гены различаются, то особь называется гетерозиготной (по данному гену), а если не различаются, то гомозиготной. При инбридинге родители являются родственниками и поэтому имеют много одинаковых генов, в результате чего гомозиготность увеличивается с каждым поколением.
56.Пренатальная диагностика наследственных заболеваний человека. Методы пренатальной диагоностики и их возможности.
В связи с отсутствием в настоящее время действенных методов лечения, тяжелым поражением здоровья при многих наследственных заболеваниях их ранняя диагностика дает возможность предупредить появление потомства с наследственным нарушением путем прерывания беременности, а иногда начать лечение сразу после рождения или даже в пренатальном периоде.
Амниоцентез, с помощью которого на 15—16-й неделе беременности получают амниотическую жидкость, содержащую продукты жизнедеятельности плода и клетки его кожи и слизистых. Забираемый при амниоцентезе материал используют для биохимических, цитогенетических и молекулярно-биологических исследований. Цитогенетическими методами определяют пол плода и выявляют хромосомные и геномные мутации. Изучение амниотической жидкости и клеток плода с помощью биохимических методов позволяет обнаружить дефект белковых продуктов генов, однако не дает возможности определять локализацию мутаций в структурной или регуляторной части генома. Важную роль в выявлении наследственных заболеваний и точной локализации повреждения наследственного материала плода играет использование ДНК-зондов.
В настоящее время с помощью амниоцентеза диагностируются все хромосомные аномалии, свыше 60 наследственных болезней обмена веществ, несовместимость матери и плода по эритроцитарным антигенам.
С начала 80-х гг. XX в. стало возможным использование для целей медикогенетического диагностирования материала биопсии ворсин хориона. В отличие от амниоцентеза это исследование проводят в первой трети беременности, что позволяет при наличии показаний прерывать ее в более ранние сроки.
Для диагностики таких заболеваний, как гемоглобинопатия, используют пункцию сосудов плода с получением клеток его крови.
Методы фетоскопии и ультразвуковых исследований позволяют определять пол плода и некоторые пороки его развития путем непосредственного наблюдения.
Пренатальная диагностика должна проводиться до 20—22-й недели беременности, когда плод еще нежизнеспособен после ее прерывания. Прерывание беременности в более поздние сроки может привести к рождению живого ребенка и быть опасным для организма матери. Прерывание беременности всегда проводится только с согласия родителей.
Так как многие методы пренатального обследования плода не являются абсолютно безвредными, а кроме того, они трудоемки и дорогостоящи, показания к такому обследованию должны быть обоснованы.
Пренатальное обследование плода проводят в случаях: 1) обнаружения структурных перестроек хромосом (транслокаций) у одного из родителей; 2) при наличии у родителей доминантного наследственного заболевания; 3) при наличии в семье детей с рецессивным наследственным заболеванием, что свидетельствует о гетерозиготности родителей; 4) при возрасте матери старше 35 лет, что прогрессивно повышает вероятность рождения у нее потомства с наследственной патологией; 5) при привычных выкидышах, вызывающих подозрение на несовместимость матери и плода по эритроцитарным антигенам; 6) при наличии в семье детей с врожденными пороками развития.
Благодаря разработке способов пренатальной диагностики удается сократить число рождающихся с наследственными заболеваниями.
Существуют 3 вида пренатальной диагностики.
– прямые (исследование плода)
– непрямые (исследование матери)
– дополнительные (УЗИ).
Прямые исследования – ранний метод. Используется плодный материал, ставят диагноз до 20 дней. Используются амниоцентез, хондроцентез – взятие крови из пупочных сосудов плода, плацентобиопсия – исследование материала плаценты, хориобиопсия – взятие ворсинок в слизистой матки, определение питания на 8-10 недели беременности. Результат изучается в течение дня, изучается риск абортов и диагностических ошибок. Амниоцентез был предложен немецким гинекологом в 19 веке. На 16 – 17 неделях делается абдоминальный прокол и забирается 10 – 20 мл амниотической жидкости. Вреда не приносит, в околоплодной жидкости имеются 20% жизнеспособных клеток плода – клетки слущенного дыхательного эпителия. Вероятность ошибки 2-3% ( аномальный кариотип). При центрифугировании клетки осаждаются, затем используются различные биохимические методы. Определяется пол плода, проводят кариотипирование.
57.Закономерности наследования внеядерных генов. Наследственные митохондриальные болезни.
58.Общие подходы к лечению наследственных заболеваний человека.
59.Онтогенез как процесс реализации наследственной информации в определенных условиях среды. Этапы, периоды и стадии онтогенеза. Периодизация онтогенеза.
60.Характеристика и значение основных этапов эмбрионального развития: предзиготный период (прогенез), оплодотворение, дробление, гаструляция, первичный органогенез (нейруляция). Дифференцировка зародышевых листков. Образование органов и тканей.
61.Провизорные органы зародышей позвоночных (амнион, хорион, аллантоис, желточный мешок, плацента), их функции.
Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
Хорион (сероза) — самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция — дыхательный газообмен; у млекопитающих он выполняет гораздо более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, например гормонов.
Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мезодермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например в спланхноплевре желточного мешка, развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.
У млекопитающих нет запасов желтка и сохранение желточного мешка может быть связано с важными вторичными функциями. Энтодерма желточного мешка служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке.
Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя — сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
62.Особенности эмбрионального развития человека.
Эмбриональный период.
В организмах, размножающихся половым путем, начинается с момента образования зиготы, а заканчивается по-разному: выходом из яйцевых оболочек (свободный личиночный тип эмбриогенеза), выходом из зародышевых оболочек (неличиночный тип), рождением (внутриутробный, вторично личиночный). Развитие разнообразно, но можно сгруппировать по принципу отношений с окружающей средой.
Тип эмбриогенеза – совокупность признаков, которые обеспечивают развивающемуся организму связь со средой.
Свободный личиночный тип характерен для животных, откладывающих небольшие яйца с маленьким количеством желтка. Наличие личинки – особой формы, отличающейся от взрослого организма. Личинка после метаморфоза превращается во взрослую особь. У многих морских животных, у сосальщиков и других паразитов.
Неличиночный тип – яйца крупные, желтка много. Зародыши долгое время находятся под защитой яйцевых оболочек, используя запасы питательных веществ, отложенных в яйцеклетке. Акулы, скаты, круглые и плоские черви, многие насекомые и рептилии, птицы и яйцекладущие млекопитающие.
Вторично личиночный тип – яйца мелкие, из яиц выходят подвижные зародыши, способные питаться. Развитие происходит под защитой специальных образований (капсул), в случае живорождения – в организме матери. Живорождение характерно для плацентарных, тропических скорпионов, сумчатых млекопитающих и некоторых рыб и насекомых.
В эмбриогенезе человека выделяют 4 периода.
1. Начальный (1 неделя развития, до момента имплантации зародыша в слизистую матки).
2. Эмбриональный (2-8 недели).
3. Предплодный (9-12 недели).
4. Плодный (13 неделя – рождение).
В эмбриональном периоде происходят гаструляция, бластуляция, нейруляция. В предплодном имеет место интенсивный органогенез, анатомическая закладка органов. Плодный период характеризуется созданием плода под защитой плодных оболочек.
На начальном периоде имеется зигота – 1 клетка зародыша, в ней определяются отдельные участки цитоплазмы, происходят синтез ДНК, белков. Зигота обладает бисимитрическим строением. Постепенно происходит нарушение соотношения ядра и цитоплазмы, в результате происходит стимуляция процесса деления – дробления
Стадия дробления – период интенсивных клеточных делений. Размер зародыша не увеличивается, а синтетические процессы идут активно. Происходит интенсивный синтез ДНК, РНК, гистоновых и других белков. Дробление выполняет функции:
– образуется достаточное количество клеток, необходимых для формирования тканей и органов.
– перераспределение желтка и цитоплазмы между дочерними клетками. 1 и 2 борозды деления идут по меридиану, а 3 по экватору. Ближе к анимальному полюсу.
– определяется план зародыша – спинно-брюшная ось, переднее-задняя ось.
– нормализуются ядерно-цитоплазматические отношения. Количество ядер растет, объем и масса сохраняются.
Постепенно деление замедляется.
У женщины через сутки после оплодотворения во второй трети яйцевода начинается дробление. Одновременно с этим процессом зигота перемещается по яйцеводу к полости матки. Образуются клетки 2 типов: более мелкие, в последствии они дают вспомогательную ткань – трофобласт, и более крупные клетки – эмбриобласт – «зачаток зародыша», дадут начало всем клеткам организма и клеткам некоторых вспомогательных частей.
На 6-7 день после оплодотворения человеческий зародыш имеет размер 0,5 мм и состоит из 200 клеток. Он начинает прикрепляться к внутренней стенке матки, внедряется в слизистую матки, происходит имплантация. В течение суток погружается наполовину, спустя еще сутки – полностью. Затем мощно развивается трофобласт – образуются ворсинки, выросты, соприкасающиеся с кровью матери, получая питательные вещества и снабжая ими зародыша.
На 2 неделе разрастаются внезародышевые части, т.е. те части, которые образованы зародышем, но играют сначала вспомогательную роль – амнион, хорион, желточный мешок. Это провизорные органы – ценогенетические структуры, не принимающие участие в формировании взрослого организма. Клеточный материал, из которого развивается зародыш – зародышевый щиток. На ранних этапах идет подготовительная работа, развивается не сам зародыш, а части, создающие необходимые условия для существования зародыша и обеспечивающие функции дыхания, питания, выведения продуктов метаболизма, создающие жидкую среду вокруг зародыша для защиты его.
3 неделя – формируется плацента, греч. «Лепешка». Состоит из 2 частей – зародышевой и материнской. Зародышевая – трофобласт и некоторые другие ткани (хорион – греч. «оболочка, послед»). Материнская – сильно видоизмененная слизистая оболочка матки. В ней разрушаются сосуды, разрыхляется соединительная ткань, разрушается и эпителий. Ворсинки хориона «купаются» в материнской крови. Площадь плацентарного сплетения 5 квадратных метров, а общая длина ворсинок хориона – 5 км. Материнский и зародышевый организмы не имеют общего кровотока, кровь не смешивается. Питательные вещества идут через стенки хориона. У 3 недельного зародыша появляются пупочные сосуды, врастающие в стенки хориона и выполняющие функции. Питания.
4 неделя. Размеры зародыша вместе с хорионом 5-7 мм. Начинается новый этап. Тело зародыша обособляется от внезародышевых частей. Зародыш приподнимается над амниотической жидкостью, с которой он связан затем только пупочными сосудами. В ходе эмбрионального развития у человека рано возникает желточный мешок – первый орган кроветворения, выполняющий запасание и переработку желтка, первый орган дыхания, питания. Первичные половые клетки начинают формироваться в желточном мешке. Имеется слепо замкнутый с 2 сторон кишечник. Печень – орган кроветворения. Бьется сердце. К концу 4 недели есть зачаток дыхательной системы. Размеры до 30мм.
Кишечник растет в длину, в выпрямленном состоянии не помещается и начинает изгибаться. К концу 4 недели появляются лопаточки на боках. В них врастают нервы и мышцы – будущие руки и ноги. К концу недели существует дифференциация на части, к 5 неделе по бокам задней части головы и шеи впячиваются участки зародыша – образуются 4 пары жаберных щелей, изнутри выпячиваются части передней кишки, формируют 4 жаберных кармана. Между жаберными щелями и жаберными карманами нет связи. Из 1 пары жаберных щелей формируется среднее ухо. Из остальных – щитовидная и зобная железы.
С 4 недели начинает формироваться нервная система. Образование нервной трубки (нервна пластинка – нервный желобок – нервная трубка). На переднем конце нервной пластинки возникают 3 мозговых пузыря, на 6 неделе уже имеются 5 мозговых пузырей, которые соответствуют отделам мозга, появляются слуховые пузырьки, глазные бокалы, обонятельные ямки. Происходит дифференциация мезодермы. Образуется хвост (34 день) до 10 мм.
На 2 месяце закладываются первичные половые железы, куда мигрируют первичные половые клетки из желточного мешка.
На 8 неделе происходит быстрое развитие амниотической оболочки и накопление жидкости.
§
Рост организма может быть определенным и неопределенным.
Определенный рост – рост прекращается к определенному времени: птицы, насекомые, млекопитающие, человек.
Неопределенный рост – постоянный рост: растения, рыбы, земноводные.
Возрастная периодизация жизни человека (1965).
| пол | возраст | название периода |
| М, Ж | 1-10 дней | Новорожденный, неонатальный |
| М, Ж | 10 дней – 1 год | Грудной |
| М, Ж | 1 – 3 года | Раннее детство |
| М, Ж | 4 – 7 лет | Первое детство |
| Ж | 8 – 11 лет | Второе детство |
| М | 8 – 12 лет | |
| Ж | 12 – 15 лет | Подростковый возраст |
| М | 13 – 16 лет | |
| Ж | 16 – 20 лет | Юношеский возраст |
| М | 17 – 21 год | |
| Ж | 21 – 35 лет | Зрелый возраст ( 1 период) |
| М | 22 – 35 лет | |
| Ж | 36 – 55 лет | Зрелый возраст ( 2 период) |
| М | 36 – 60 лет | |
| Ж | 56 – 74 года | Пожилой возраст |
| М | 61 – 74 года | |
| М, Ж | 75 – 90 лет | Старческий |
| М, Ж | Более 90лет | Долгожители |
Период новорожденности. Приспособление ребенка к новым для него условиям внешней среды. Начинают функционировать система внешнего дыхания, желудочно-кишечный тракт, меняется характер кровоснабжения, со стороны центральной нервной системы преобладают процессы торможения. Температура тела неустойчива в первые дни ( /-2,5 градуса). К концу недели катаболическая фаза сменяется анаболической. Начинается рост тела, улучшаются показатели ферментативных и других систем.
Грудной возраст. Значительное усиление обменной активности при выраженной незрелости систем. Увеличение числа клеток. Размеров органов. Формирование и специализация органов, увеличение массы тела, изменение состава крови: замена фетального гемоглобина на более зрелый гемоглобин. Органы пищеварения, дыхания имеют очень ограниченные возможности. Быстро происходит созревание центральной нервной системы и периферической нервной системы. Приобретение и накопление условно-рефлекторных связей. К концу года повышается психомоторное развитие ребенка.
Изменение длины тела.
| возраст | длина |
| Новорожденный | 46 – 56 |
| За 1 год | 25 см |
| За 2 год | 12 – 13 см |
| За 3 год | 12 – 13 см |
| 4-7 лет (ежегодно) | 5 – 7 см |
| Второе детство (ежегодно) | 4 – 5 см |
| Период полового созревания (ежегодно) | 7 – 8 см |
Масса тела новорожденного. У детей до года увеличение длины тела на 1 см сопровождается увеличением массы на 280 – 320 г. У детей более старшего возраста определяется по формулам.
Поверхность тела на 1 кг его массы у новорожденного в 3 раза больше, чем у взрослого.
Масса мозга новорожденного 330 – 430 г. К концу первого года жизни увеличивается в 2 – 2,5 раза, к концу второго года в 3 раза. После 7 лет увеличение мозга замедляется, а к 20 – 29 годам достигает максимума.(1355г у мужчин и 1220 г у женщин). Количество борозд и извилин у новорожденного такое же, как и у взрослого, но их дифференцировка и развитие выражены слабо. Дифференцировка нервных клеток идет в первые 5 – 6 месяцев и заканчивается в основном к 8 годам. Кора больших полушарий по строению похожа на кору больших полушарий взрослого человека. Совершенствуется двигательная область коры и появляется способность к тонким коордиционным движениям. Активное развитие спинного мозга, к 2 годам как у взрослого.
К 4 годам формируется нижний носовой ход, способствующий согреванию и очищению воздуха.
Жизненная емкость легких, количество альвеол к концу дошкольного периода, как у взрослого. Жизненная емкость легких в 4 года – 1100 мл, в 6 лет -1200 мл.
Масса сердца к 5 годам увеличивается в 4 раза, к 6 годам в 11 раз по сравнению с исходной. Происходит окончательная дифференцировка.
К5-7 лет прорезаются постоянные большие коренные зубы (первые – слева).
Органы чувств. К 5 – 7 годам хорошо развиты все виды чувствительности кожи. Продолжается совершенствование иммунной системы. Процесс иммунизации заканчивается до половой зрелости.
Сроки полового созревания для девочек -12-16 лет, для мальчиков – 13 – 18 лет. В этот период наблюдается быстрое скачкообразное увеличение размеров тела, жизненной емкости легких, мышечной силы и работоспособности. Это пубертатный период. В это время максимальная скорость роста у мальчиков – до 10 см в год. У девочек немного меньше. Пубертатный период у девочек на 2 года меньше. У многих детей наблюдается регулярное сезонное увеличение длины тела (весной быстрее, осенью – медленнее). С изменением роста совладает изменение скелетных размеров, мышечной массы, рост печени, почек.
Не все ткани и системы органов растут одинаково. Выделяют 4 типа.
1. Общий тип в целом тело. Мышцы, скелет, органы дыхания, печень. Повторяют ход кривой роста длины тела. Имеются 2 пика – в первый год жизни и в пубертатный период
2. Мозговой и головной тип. Головной, спинной мозг, глаза, размеры головы. Развиваются раньше. Интенсивный рост: к 10-12 годам достигают размеров взрослого человека.
3. Лимфоидный тип. Тимус, лимфатические узлы, лимфатическая ткань кишечника, селезенки, миндалин. Интенсивного развития достигают к 11 – 12 годам (большего размера, чем у взрослых). У людей старшее 35 – 40 лет происходит редукция лимфатических сосудов. Некоторые лимфатические сосуды запустевают.
4. Репродуктивный тип. Предстательная железа, семенные пузырьки, яичники, фаллопиевы трубы. Почти не растут до периода полового созревания, а затем быстро достигают размеров взрослого человека.
На 34 неделе эмбрионального развития у плода закладывается подкожная жировая клетчатка. Продолжается до 6 -7 месяца после рождения. До 6 – 7 лет отсутствует, потом начинается вновь. У мальчиков на конечностях падает, у девочек на конечностях приостанавливается, но не уменьшается. Продолжается на теле до 16 лет.
Дорепродуктивный период направлен на подготовку к размножению. Те организмы, что дожили до половой зрелости, участвуют в эволюционном процессе.
Репродуктивный период у женщин до 45 – 55 лет характерная особенность – преобладание скорости воспроизведения в начале периода, которая падает к концу периода. Позвоночный столб растет до 30 – 33 лет, в 30 – 50 лет длина позвоночника постоянна. В 20 – 60 лет происходит увеличение большей части головы и лица на 2 – 4%.
На рост и развитие детей сильное влияние оказывает социально – экономическое развитие. Деи из более обеспеченных семей быстрее растут. Последние 100 – 120 лет произошло увеличение роста детей (характерно для стран Европы, Японии) – акселерация. Грудные дети чаще увеличивают вес. На 1 год раньше прорезываются зубы. Половое созревание раньше на 2 года. Юноши стали выше на 12 – 14 см. Темп акселерации замедляется. Есть отрицательные моменты акселерации: у таких детей часты неврозы, нарушения высшей нервной деятельности. Не очень благоприятные отношения между ростом. Весом и размерами сердца.
Четких границ между репродуктивным и пострепродуктивным периодами нет. Переход постепенен. Он характеризуется постепенным угасанием функций организма и отдельных органов.
Старение как закономерный этап онтогенеза. Проявления старения на молекулярно-генетическом, клеточном, тканевом, органном и организменном уровнях.
Старение– многоэтапный процесс, включающий влияние факторов внешней и внутренней среды. Старение общебиологическая закономерность «увядания» организма, свойственная всем живым существам.
Старость – естественный этап онтогенеза, заканчивающийся смертью. Старость – следствие старения.
Карл Маркс: « Мы должны рассматривать старение и смерть как естественный завершающий этап индивидуального развития. Жить – значит умирать».
У разных биологических видов продолжительность жизни различна (даже у видов, занимающих сходные экологические ниши). Поэтому продолжительность жизни является видовой биологической индивидуальностью, сложившейся в ходе эволюционного процесса, контролируемой генетически. В пределах вида возможны отклонения до 50% средней продолжительности жизни в обе стороны.
Кенгуру – 5 лет.
Бобр – 50 лет.
Тигр – 40 – 50 лет.
Осел – 47 лет.
Индийский слон – 70 – 77 лет.
У человека средняя продолжительность жизни мужчин – 58 – 62 года, у женщин – 74 года (Воронеж).
Хронологический возраст – количество прожитых лет человеком по документам.
Биологический возраст – показывает, на сколько выглядит человек, (степень окостенения, зубная зрелость, степень развития половых органов).
Признаки старения.
Старение происходит на всех уровнях
1. во внешних признаках
изменяется осанка, форма тела
появляется седина
теряется эластичность кожи, что приводит к появлению морщин
ослабляется зрение и слух
ухудшается память
психомоторная реакция начинает замедляться к 25 – 30 годам, память – к 30 годам, способность к обучению – к 20 годам.
2. на уровне органов
уменьшается жизненная емкость легких
повышается артериальное давление
развивается атеросклероз
происходит инволюция половых желез
уменьшается продукция половых желез и гормонов щитовидной железы
падает основный обмен
уменьшается работа желудочно–кишечного тракта.
3. на уровне клеток
падает количество воды
уменьшается активность окислительного фосфорилирования в ферментных системах
уменьшается репликация ДНК
падает активность синтеза РНК
увеличивается количество генных и хромосомных мутаций из-за снижения эффективности процесса репарации.
Старость и болезни – единый патологический процесс. Например, наследственные синдромы преждевременного старения (прогерии). Симптомы: поседение, морщинистость кожи, облысение, атеросклероз, инфаркт миокарда, гиперхолестерин и другие симптомы. Могут проявляться в детском возрасте. В 3 – 5 лет имеют вид старческий вид, к 7 годам умирают от старости. Средняя продолжительность жизни таких больных – 12 лет, иногда доживали до 30 лет (максимум – 39 лет).
Прогерии– наследственные заболевания, при которых получают резкое развитие один или несколько симптомов есте6ственного старения.
Гипотезы старения.
Существуют более300 различных гипотез.
1. Энергетическая. Автор – Рубнер- 1908 год. Каждый вид имеет определенный энергетический фон, распространив который организм стареет и умирает.
2. Гормональная теория. Причина старения – снижение синтеза половых гормонов.
3. Интоксикационная. Автор – Мечников. Самоотравление организма, в основном вызванное гниением в толстом кишечнике.
4. Перенапряжение ЦНС. Автор – Павлов. Нервные потрясения и стрессы приводят к старению
5. Соединительнотканная теория. Автор – Богомолец. В результате нарушения межтканевых взаимодействий наступает старение.
6. Генетические теории старения – наиболее молодые. Первично возникающие изменения генетического аппарата клеток приводят к повышению количества мутаций, падению скорости синтеза ДНК и старению.
7. Программная теория. Основана на том, что в организме функционируют особые часы. Которые запускают механизмы возрастных изменений.
8. Теория Хейфлига – 1965 год. Количество митозов ограничено. Клетки организма – 50 – 60 потомки зиготы. Более 50 – 60 раз клетки не делятся. Рассматривал эту теорию на фибробластах.
Геронтология – наука о старости, изучающая основные закономерности старения на всех уровнях организации от молекулярного до организменного.
Гериатрия– наука, изучающая особенности развития, течения, предупреждения заболеваний у людей преклонного возраста.
Задача геронтологии – качественное и количественное продление жизни человека.
65.Гипотезы, объясняющие механизмы старения. Влияние генетических факторов, условий и образа жизни на процесс старения.
66.Смерть как биологическое явление. Клиническая и биологическая смерть. Реанимация. Проблемы долголетия. Понятие о геронтологии и гериатрии.
Старение– многоэтапный процесс, включающий влияние факторов внешней и внутренней среды. Старение общебиологическая закономерность «увядания» организма, свойственная всем живым существам.
Старость – естественный этап онтогенеза, заканчивающийся смертью. Старость – следствие старения.
Карл Маркс: « Мы должны рассматривать старение и смерть как естественный завершающий этап индивидуального развития. Жить – значит умирать».
У разных биологических видов продолжительность жизни различна (даже у видов, занимающих сходные экологические ниши). Поэтому продолжительность жизни является видовой биологической индивидуальностью, сложившейся в ходе эволюционного процесса, контролируемой генетически. В пределах вида возможны отклонения до 50% средней продолжительности жизни в обе стороны.
Кенгуру – 5 лет.
Бобр – 50 лет.
Тигр – 40 – 50 лет.
Осел – 47 лет.
Индийский слон – 70 – 77 лет.
У человека средняя продолжительность жизни мужчин – 58 – 62 года, у женщин – 74 года (Воронеж).
Хронологический возраст – количество прожитых лет человеком по документам.
Биологический возраст – показывает, на сколько выглядит человек, (степень окостенения, зубная зрелость, степень развития половых органов).
Признаки старения.
Старение происходит на всех уровнях
1. во внешних признаках
1.1 изменяется осанка, форма тела
1.2 появляется седина
1.3 теряется эластичность кожи, что приводит к появлению морщин
1.4 ослабляется зрение и слух
1.5 ухудшается память
1.6 психомоторная реакция начинает замедляться к 25 – 30 годам, память – к 30 годам, способность к обучению – к 20 годам.
2. на уровне органов
2.1 уменьшается жизненная емкость легких
2.2 повышается артериальное давление
2.3 развивается атеросклероз
2.4 происходит инволюция половых желез
2.5 уменьшается продукция половых желез и гормонов щитовидной железы
2.6 падает основный обмен
2.7 уменьшается работа желудочно–кишечного тракта.
3. на уровне клеток
3.1 падает количество воды
3.2 уменьшается активность окислительного фосфорилирования в ферментных системах
3.3 уменьшается репликация ДНК
3.4 падает активность синтеза РНК
3.5 увеличивается количество генных и хромосомных мутаций из-за снижения эффективности процесса репарации.
Старость и болезни – единый патологический процесс. Например, наследственные синдромы преждевременного старения (прогерии). Симптомы: поседение, морщинистость кожи, облысение, атеросклероз, инфаркт миокарда, гиперхолестерин и другие симптомы. Могут проявляться в детском возрасте. В 3 – 5 лет имеют вид старческий вид, к 7 годам умирают от старости. Средняя продолжительность жизни таких больных – 12 лет, иногда доживали до 30 лет (максимум – 39 лет).
Прогерии– наследственные заболевания, при которых получают резкое развитие один или несколько симптомов есте6ственного старения.
Гипотезы старения.
Существуют более300 различных гипотез.
· Энергетическая. Автор – Рубнер- 1908 год. Каждый вид имеет определенный энергетический фон, распространив который организм стареет и умирает.
· Гормональная теория. Причина старения – снижение синтеза половых гормонов.
· Интоксикационная. Автор – Мечников. Самоотравление организма, в основном вызванное гниением в толстом кишечнике.
· Перенапряжение ЦНС. Автор – Павлов. Нервные потрясения и стрессы приводят к старению
· Соединительнотканная теория. Автор – Богомолец. В результате нарушения межтканевых взаимодействий наступает старение.
· Генетические теории старения – наиболее молодые. Первично возникающие изменения генетического аппарата клеток приводят к повышению количества мутаций, падению скорости синтеза ДНК и старению.
· Программная теория. Основана на том, что в организме функционируют особые часы. Которые запускают механизмы возрастных изменений.
· Теория Хейфлига – 1965 год. Количество митозов ограничено. Клетки организма – 50 – 60 потомки зиготы. Более 50 – 60 раз клетки не делятся. Рассматривал эту теорию на фибробластах.
Геронтология – наука о старости, изучающая основные закономерности старения на всех уровнях организации от молекулярного до организменного.
Гериатрия– наука, изучающая особенности развития, течения, предупреждения заболеваний у людей преклонного возраста.
Задача геронтологии – качественное и количественное продление жизни человека.
СТАРОСТЬ И СТАРЕНИЕ.
§
Рассмотрим кратко изменения органов и функциональных систем животных и человека, которые становятся заметными и нарастают по завершении активного репродуктивного периода онтогенеза.
Как правило, после 40—50 лет у человека возникают стойкие внешние проявления старения, в частности кожных покровов. Появляются морщины, образующиеся из-за потери подкожной жировой ткани, пигментные пятна, бородавки. Кожа становится сухой и шершавой в связи с уменьшением количества потовых желез, теряется ее эластичность, она становится дряблой.
Общее направление этих изменений для сердечно-сосудистой, дыхательной систем, а также основного обмена, отражающего состояние энергетических процессов в организме, иллюстрирует рис. 8.29.
Признаки старения сердечно-сосудистой системы становятся заметными обычно в возрасте после 40 лет. Закономерные изменения наблюдаются в стенках сосудов: в них откладываются липиды, прежде всего холестерин, что наряду с другими структурными превращениями снижает эластичность и искажает ответы на различные стимулы, регулирующие кровообращение. Типичным является разрастание в стенках сосудов и сердца соединительной ткани, замещающей рабочую мышечную ткань. В результате снижается эффективность работы сердца. В итоге нарушается кровоснабжение тканей и органов. Так, кровоток по сосудам головного мозга 75-летнего человека по сравнению с 30-летним уменьшен на
В основе функциональных расстройств дыхательной системы лежит разрушение межальвеолярных перегородок, что сокращает дыхательную поверхность, разрастание в легких соединительной ткани, снижает эффективность аэрогематического обмена кислорода. В итоге с возрастом падает жизненная емкость легких, которая к 75 годам достигает всего 56% от уровня в возрасте 30 лет.
Легко заметным изменением в системе пищеварения является потеря зубов. Падает эффективность функционирования пищеварительных желез, нарушения двигательной (моторной) функции кишечника нередко приводят к привычным запорам.
В процессе старения страдает функция мочевыделительной системы, снижается интенсивность фильтрации в почечных клубочках (на 31% в 75-летнем возрасте по сравнению с 30-летним), так же как и обратное всасывание веществ из фильтрата в почечных канальцах. Ухудшение функции мочевыделения объясняется гибелью с возрастом значительного количества нефронов (до 44% от уровня 30-летнего возраста), представляющих собой структурно-функциональные единицы почек.
Специального внимания заслуживают изменения в процессе старения со стороны мышечной системы и скелета. Снижается сила сокращений поперечно-полосатой мускулатуры, быстрее развивается утомление, наблюдается атрофия мышц. Характерная для стареющих людей перестройка костей заключается в разрежении их вещества (старческий остеопороз), что приводит к снижению прочности.
В процессе старения организма существенные изменения происходят в репродуктивной системе. При этом они затрагивают обе основные функции главных органов названной системы — половых желез: выработку гамет и образование половых гормонов. У женщин овогенез прекращается по достижении ими менопаузы. Образование функционально полноценных сперматозоидов в мужском организме возможно, по-видимому, даже в преклонном возрасте.
Изменение гормонального профиля людей в связи с угасанием репродуктивной функции носит сложный характер. Распространенным является мнение о прогрессивном снижении с возрастом концентрации у мужчин тестостерона, а у женщин эстрадиола и прогестерона — главных мужского и женских половых гормонов. Напомним, что оба типа гормонов образуются организмами обоих полов, только в разном количестве. Указанные сдвиги сопровождаются повышением секреции эстрадиола и прогестерона у мужчин и тестостерона у женщин. Вместе с тем содержание фолликулостимулирующего гормона у 80—90-летних женщин выше в 14 раз, а лютеинизирующего гормона — в 5 раз, чем у 20—30-летних. Резко нарушено у старых людей соотношение названных гормонов гипофиза, что является важной причиной нарушения репродуктивной функции в целом. Картина усложняется также тем, что в процессе старения изменяется ответ ткани на половые гормоны в связи с сокращением количества клеточных рецепторов к ним.
Изменения в процессе старения функций эндокринной системы носят объективно сложный характер. В качестве примера рассмотрим изменения в стареющем организме функции щитовидной железы. Обнаружено, что к старости падает содержание в крови трииодтиронина и тироксина, в связи с изменением белков плазмы крови ухудшается перенос гормонов к тканям, в клетках уменьшается количество рецепторов, узнающих гормоны, а чувствительность рецепторов повышается. Вместе с тем в крови сохраняется достаточно высокая концентрация тиреотропного гормона гипофиза; чувствительность к нему клеток щитовидной железы возрастает. Из приведенной картины видно, что в отдельных звеньях цепи регуляции жизненно важных функций гормонами щитовидной железы возрастные изменения не одинаковы по масштабу, а иногда и разнонаправлены. В таком случае важен общий результат, степень выраженности которого подвержена индивидуальным колебаниям.
Наряду с эндокринной системой нервной системе принадлежит важнейшая роль в регуляции, координации и интеграции разнообразных проявлений жизнедеятельности. Изменения нервной системы в процессе старения включают нарастающую гибель нейронов, масштабы которой колеблются от отдела к отделу и составляют от 15—20 до 70—75%. С другой стороны, сохранившиеся клетки увеличивают свои размеры, образуют дополнительные разветвления окончаний отростков в тканях-мишенях, что имеет приспособительное, заместительное значение. Общий итог, однако, заключается в том, что после 40—50 лет в тканях образуется недостаток нервных влияний, что приводит к нарушению регуляций функций. Это справедливо, к примеру, в отношении иннервации сердца и сосудов, а также поперечно-полосатой скелетной мускулатуры симпатической нервной системой.
Функциональные нарушения в процессе старения в центральной нервной системе зависят как от уменьшения количества нервных клеток, так и от снижения синтеза медиаторов, ослабления связей между отдельньми мозговыми структурами, уменьшения скорости проведения нервных импульсов по волокнам и через синапсы. В стареющем организме с большим трудом вырабатываются условные рефлексы, приобретаются новые навыки. Изменения ряда функций носят более сложный характер. Так, интеллектуальные способности людей в возрасте 55—70 лет сохраняются такими же, какими они были в 20—30-летнем возрасте, а в возрасте 55—60 лет наблюдается второй пик творческой деятельности.
Одна из черт процесса старения заключается в снижении надежности механизмов регуляции, направленных на поддержание постоянства жизненно важных параметров внутренней среды организма — гомеостаза. Причину этого видят в функциональных изменениях стареющего гипоталамуса, с которым связывают действия своеобразных биологических часов старения всего организма. К гипоталамической области головного мозга человека относятся 32 пары ядер, участвующих в регуляции важнейших вегетативных функций. В целом процесс старения гипоталамических структур характеризуется неравномерностью и разнонаправленностью, что типично для других областей нервной системы.
В старости наблюдается снижение функций практически всех органов чувств. Уменьшается способность глаза к аккомодации, так как слабеют глазодвигательные мышцы и изменяется вещество хрусталика. Это ведет к старческой дальнозоркости. У старых людей хрусталик нередко теряет прозрачность — развивается катаракта. Падает острота зрения. Чувствительность органа слуха снижается, причем особенно к высокочастотным звуковым колебаниям. Старый человек хуже различает запах и вкус. К 75 годам люди сохраняют примерно 36% от количества вкусовых луковиц, которое имеется в возрасте 30 лет. У них нарушается чувство равновесия.
С возрастом заметно изменяются функции иммунной системы. За гуморальный иммунитет ответственны В-лимфоциты, вырабатывающие антитела к носителям чужеродной биологической информации — антигенам. Т-лимфоциты ответственны за клеточный иммунитет, например за отторжение трансплантата, хотя могут участвовать в реакциях гуморального иммунитета. К старости наблюдается ослабление реакций как гуморального, так и клеточного иммунитета. Становление функции иммунной системы во многом связано с активностью тимуса, которая прекращается в связи с инволюцией железы по достижении половозрелого возраста (рис. 8.30). Нарушение естественного развития иммунной системы в эксперименте путем удаления вилочковой железы (например, у мышей) сокращает продолжительность жизни животных. Напротив, пересадка тимуса и костного мозга от молодой мыши к старой приводит к омоложению иммунной системы 19-месячной мыши до уровня 4-месячной. Некоторые такие животные жили на Уз дольше своего обычного срока.
В стареющем организме клетки иммунной системы ошибочно вырабатывают антитела против собственных клеток и белков. Таким образом, старение сопровождается нарастанием аутоиммунных реакций.
Изменения в процессе старения органов кроветворной системы заключаются в снижении продукции эритроцитов. С возрастом способность костного мозга к восстановлению массы эритроцитов падает почти в б раз. Восполнение утраченных вследствие потери 30—40% общего объема крови эритроцитов происходит у 2-месячных крыс по 4,8%, а у 25-месячных —по 0,8% в 1 сут (рис. 8.31).
Из изложенного в данном разделе следует, что процесс старения распространяется практически на все структуры и функции организма. При этом некоторые показатели (упругость кожи, поседение волос) изменяются с возрастом довольно монотонно. Другие (артериальное давление, жизненная емкость легких) изменения носят пороговый характер, начиная регистрироваться примерно в возрасте 35—45 лет.
§
Молекулярные и клеточные проявления старения многообразны. Они заключаются в изменении показателей потоков информации и энергии, состояния ультраструктур дифференцированных клеток, снижении интенсивности клеточной пролиферации.
Напомним, что функционирование ДНК, заключающей в себе биологическую информацию, связано с ее репродукцией, транскрипцией, репарацией. Учитывая возможную роль ошибок в молекулах ДНК, в нарушении клеточных функций в процессе старения изучали эффективность механизмов репарации повреждений молекулярной структуры ДНК (согласно расчетным данным, ежесекундно в геноме возникает минимум одно повреждение) в разном возрасте, а также корреляцию между интенсивностью этого процесса и продолжительностью жизни. В обоих случаях были получены противоречивые результаты. С одной стороны, уровень репарации повреждения ДНК УФ-облучением эмбриональных фибробластов трех линий мышей оказался пропорциональным средней продолжительности их жизни (900, 600 и 300 сут). С другой — репарация ДНК после УФ-облучения не различалась в культурах фибробластов в коже людей в диапазоне возрастов от 0 (новорожденные) до 88 лет. Общее заключение сводится к тому, что интенсивность молекулярной репарации ДНК меняется с возрастом в некоторых типах клеток, но не это является главной причиной клеточного старения.
В дифференцированных клетках млекопитающих животных старение сопровождается в целом снижением транскрипционной активности. Так, у мышей интенсивность синтеза РНК в ядрах печеночных и нервных клеток между 12-м и 30-м месяцами жизни падает на 50%. Изменение синтеза относится не только к рРНК, не кодирующим структуру белков, но и к мРНК. В сравнении с активным репродуктивным периодом жизни в стареющем организме действительно наблюдается исчезновение в клетках определенных типов мРНК, правда, в это же время регистрируется появление некоторых типов мРНК, не образующихся ранее. Таким образом, речь идет о частичной смене биологической информации, используемой клеткой в разном возрасте.
Скорость снижения транскрипционной активности ДНК в постмитотических высокодифференцированных клетках, к примеру нервных, зависит от условий их существования в течение жизни, в частности от напряженности их функционирования. Так, одна и та же функциональная нагрузка, распределенная между меньшим количеством клеток, приводит к более раннему падению уровня транскрипции в их ядрах. Было также показано, что параллельно изменению скорости снижения транскрипции сокращается максимально достигаемая животными продолжительность жизни. ДНК эукариотических клеток находится в комплексе с белками — гистоновыми и негистоновыми, образуя вместе с ними хроматин ядер. Предполагают, что регуляция транскрипции информации с ДНК происходит путем изменения ДНК-белковых связей в хроматине. С возрастом такие связи становятся менее подвижными, отмечается снижение содержания в хроматине негистоновых белков.
Изменение трансляции в процессе старения изучают по содержанию рРНК (показатель общей белокобразующей способности клетки), мРНК (набор образуемых белков), активности аминоацил-тРНК-синтетаз (ферменты активации аминокислот). Оказалось, что в возрасте от 12 до 70 лет у людей утрачивается до половины генов рРНК, относящихся, как известно, к умеренно повторяющимся нуклеотидным последовательностям, которые продублированы в геноме более 300 раз. Сохраняющееся число генов, по-видимому, способно обеспечить образование требуемого количества рРНК. Интенсивность белкового синтеза в целом снижается в зрелом возрасте.
О возрастных изменениях набора образуемых белков судят по содержанию в клетках различных ферментов. Полученные данные трудно оценить однозначно, так как обнаруженные отклонения даже в группах ферментов, сходных по функции, нередко разнонаправленны. Вместе с тем активность ферментов, ответственных за окисление, изменяется в стареющем организме однонаправленно: она снижается.
Немаловажное значение при старении имеют изменения энергетики организма, в частности обратная связь между продолжительностью жизни животных различных видов и удельной скоростью обмена веществ отмечена давно. Существует особое понятие энергетического жизненного потенциала, отражающего общее количество расходуемой за жизнь энергии. Его величина для млекопитающих (кроме приматов) составляет примерно 924 кДж/г, большинства приматов — 1924, лемура, обезьяны-капуцина и человека — 3280 кДж/г массы тела. Изменения потока энергии в процессе старения состоят в снижении количества митохондрий в клетках, а также падении эффективности их функционирования. Так, у взрослых крыс количество кислорода, потребляемое на 1 мг белка митохондрий, более чем в 1,5 раза выше, чем у старых животных. Важным свойством стареющего организма является смещение в процессах энергообеспечения функций соотношения между тканевым дыханием и гликолизом (бескислородный путь образования АТФ) в пользу последнего.
Изменения в процессе старения ультраструктуры клеток затрагивают практически все органеллы как общего, так и специального значения. Одновременно может происходить накопление необычных веществ, иногда структурно оформленных (липофусцин). Наиболее заметна возрастная перестройка постмитотических высокоспециализированных клеток—нейронов, кардиомиоцитов. Для стареющих нервных клеток, например, типично обеднение цитоплазмы мембранами, сокращение объема шероховатой эндоплазматической сети, увеличение содержания в клеточных телах микрофибрилл, что, возможно, связано с нарушением транспорта веществ по отросткам.
В отростках мотонейронов старых крыс скорость транспорта составляет примерно 200, тогда как у зрелых животных —320 мм/сут. Параллельно наблюдается снижение интенсивности синтеза белка и РНК. Отмечается замедление проведения нервного импульса, а в некоторых типах нервных клеток — уменьшение количества образуемого медиатора. Наиболее типичной чертой старения нервных клеток млекопитающих и человека является нарастающее накопление с возрастом в цитоплазме пигмента липофусцина. У 60-летних людей благодаря увеличению содержания пигмента доля цитоплазмы снижается в 1,3, ay 80-летних —в 2 раза в сравнении с 40-летними. Липофусцин часто называют пигментом изнашивания, т.е. балластом. Противоположная точка зрения приписывает липофусцину роль внутриклеточного депо кислорода.
Возрастное накопление липофусцина распространяется кроме нервной системы на сердечную и скелетную мускулатуру. Сдерживание роста содержания пигмента в клетках плодовых мух путем ограничения летательной активности сочеталось с двукратным увеличением средней продолжительности жизни.
Еще один пример изменения в ходе старения специальных органелл касается микрофибрилл в клетках сердечной мышцы, в отношении которых начинают преобладать деструктивные процессы.
На определенном этапе прогрессивная эволюция жизни на земле оказалась связанной с переходом к более эффективному аэробному типу энергообеспечения процессов жизнедеятельности.Не следует, однако, забывать, что использование клетками кислорода приводит к образованию свободных радикалов (О2, ОН, Н2О2), которые в силу чрезвычайной реакционноспособности могут вызывать быстрые разрушения биологических структур (мембран, макромолекул). Неблагоприятным эффектам свободных радикалов в клетках противостоят закрепленные процессом эволюции природные антиоксидантные механизмы. К ним принадлежат ферменты, разрушающие пероксиды. В процессе старения действенность механизмов, нейтрализующих свободные радикалы и пероксиды, снижается. Свободные радикалы способны нарушить любое звено молекулярной организации клетки. Сказанное делает их универсальным фактором старения на молекулярном и субклеточном уровне вне зависимости от вида клетки.
С начала 60-х гг. появились новые взгляды на значение для старения и продолжительности жизни закономерностей клеточной пролиферации. На основании подсчета числа делений фибробластов, высеваемых в культуру ткани от эмбриона человека и от людей в возрастете лет и выше, было сделано заключение о пределе клеточных делений (лимит Хейфлика), которому соответствует видовая длительность жизни. Показано, что фибробласты мыши способны удваивать свою численность 14—28 раз, цыпленка — 15—35, человека — 40—60, черепахи — 72— 114 раз. Проверка результатов, о которых идет речь, выявила, что представление об ограниченности числа клеточных делений в индивидуальном развитии является неточным.
Вместе с тем сохраняют свое значение классические представления, уходящие корнями в XIX столетие, утверждающие, что старение и естественное его следствие — смерть — это своеобразная плата за явление клеточной дифференцировки. Выход клеток в дифференцировку для многих типов клеток означает старение и гибель в связи с утратой возможности возвращения в митотический цикл (нервные клетки, сердечная и скелетная мышцы, лейкоциты и эритроциты крови, эпителий ворсин кишечника).
§
Условия жизни относятся к категории понятий, отличающихся широтой и некоторой размытостью границ. Если для животных оно включает природно-географические, прежде всего климатические, факторы, то в отношении человека необходимо учитывать также и социально-экономические факторы.
Исследование влияний условий жизни на процесс старения про-„водят несколькими путями. Во-первых, путем изучения скорости соответствующих изменений организмов, проживающих в разных условиях. Во-вторых, путем сопоставления значений смертности или продолжительности жизни в различающихся по условиям жизни популяциях в пределах одного или разных исторических периодов времени. Продолжительность жизни здесь выступает как обобщенный показатель жизнеспособности. В-третьих, свой вклад вносит изучение распределения по планете долгожителей.
Оценка влияния условий жизни на скорость старения требует предварительного отбора демонстративных показателей, значения которых закономерно изменяются соответственно возрасту. В исследованиях зависимости скорости старения от условий жизни, проводимых на лабораторных животных, используют следующие признаки: 1) состояние белков соединительной ткани коллагена и эластина; 2) показатели сердечной деятельности и кровообращения; 3) содержание пигмента липофусцина в клетках нервной системы и сердца; 4) показатели произвольной двигательной активности; 5) способность к обучению.
Изучали влияние на скорость старения многих условий жизни: пониженной температуры окружающей среды, измененного режима двигательной активности, воздействий ионизирующим облучением, повышенного парциального давления кислорода. В опытах на плодовых мухах, например, установлено, что увеличение в атмосфере в три раза концентрации 02 приводит к ускоренному старению, что выражается в более быстром отложении в тканях липофусцина. Параллельно отмечалось сокращение вдвое длительности жизни. В другом опыте кроликов с раннего возраста систематически и длительно подвергали повышенным двигательным нагрузкам. В описанных условиях у кроликов замедлялся ритм сердечных сокращений, снижалось артериальное давление. Направление изменений со стороны сердечно-сосудистой системы дает возможность судить о снижении скорости старения животных. Одновременно было установлено, что подопытные кролики жили в 1,5 раза дольше животных с обычной для вида двигательной активностью.
В процесс старения вовлекаются все структуры и функции организма. Важным свойством этого процесса является его гетерогенность. В соответствии с этим свойством изменения, характерные для старения, появляются в различных клетках, тканях и органах в разном календарном возрасте. С другой стороны, в любом органе стареющего организма типичные для этого процесса изменения сочетаются с изменениями приспособительными, направленными на восполнение структурных и функциональных потерь. В таких условиях невозможно точно определить скорость старения организма как целостной системы путем оценки скорости старения по изменениям отдельных показателей, будь то накопление липофусцина или повышение артериального давления. Необходим обобщенный критерий, каковым является продолжительность жизни или связанная с ней жизнеспособность.
Использование такого показателя, как продолжительность жизни, носящего статистический характер, позволяет проводить исследования непосредственно на людях, группируя условия жизни согласно их природе: социально-экономические, климатогеографические, местные, связанные, например, с минеральным составом почв и грунтовых вод или же уровнем радиации.
Влияние социально-экономических условий на длительность жизни может быть оценено путем сравнения названного показателя для одной и той же популяции (например, население страны), но в разные исторические периоды или же путем сопоставления продолжительности жизни в двух популяциях, различающихся по жизненному уровню и сосуществующих в одно и то же историческое время. Социально-экономические условия жизни населения стран Европы, Северной Америки, некоторых стран Азии и Африки претерпели существенные изменения в текущем столетии. Главным итогом этих изменений явилось повышение жизненного уровня, улучшение питания, жилищных условий, качества охраны здоровья и медицинской помощи. Соответственно значения средней продолжительности жизни в таких странах возросли к 90-м гг. XX в. более чем в два раза в сравнении со значениями показателя для начала века. К примеру, в начале века средняя продолжительность жизни жителей России составляла 32 года, тогда как в 1987 г. она достигла уровня 64 лет для мужчин и 73 для женщин.
В экономически развитых странах средняя продолжительность жизни в целом превышает 70 лет, однако во многих развивающихся странах и в настоящий исторический период она не достигает 40 лет.
Приведенные сведения указывают на очевидную зависимость длительности жизни от социально-экономических условий. Означает ли это, что изменения социально-экономических условий в сторону их улучшения замедляют скорость старения? Зависимость смертности людей от возраста показана на рис. 8.33. По ходу приведенной кривой наблюдаются три отрезка, различающихся по интенсивности и направлению изменения смертности. Во-первых, это пик смертности, вершина которого приходится на период новорожденности: примерно 2000 младенческих смертей на 100 000 рождаемых. Далее интенсивность смертности снижается, и в возрасте 9—12 лет она минимальна: 20—30 умерших на 105 детей. Начиная с возраста половой зрелости смертность монотонно возрастает, причем рост происходит по экспоненте таким образом, что после 30—35 лет удвоение значений происходит каждые 8 лет. В возрасте более 80 лет интенсивность смертности высока и по мере прибавления лет практически не меняется.
Для суждения о зависимости скорости старения людей от социально-экономических условий жизни достаточно сравнить ход кривых возрастной интенсивности смертности в популяции в начале и во второй половине XX в. Видно, что принципиальных изменений конфигурация кривой не претерпела. На всем своем протяжении она лишь сместилась в область более низких значений. Следовательно, улучшение социально-экономических условий уменьшает вероятность смерти в молодом возрасте, но не изменяет скорости старения. На неизменяемость скорости старения в конкретной популяции людей в течение XX в., несмотря на существенное повышение жизненного уровня, указывают, например, данные об абсолютном возрастном приросте интенсивности смертности мужчин Швеции с 1900 по 1980 г. (рис. 8.34). Таким образом, лучшие социально-экономические условия делают людей, притом любого возраста, лишь менее доступными действию факторов, которые в иных условиях привели бы к смертельному исходу.
Сделанное заключение хорошо согласуется с теми изменениями, которые произошли на протяжении текущего столетия в перечне главных причин смертности людей (на примере населения г. Нью-Йорка с 1900 по 1970 г.):
| Год, причина смерти 1900г. Пневмония, грипп, бронхит Туберкулез Диаррея, энтериты Болезни сердца 1930 г. Болезни сердца Пневмония, грипп, бронхит Рак Нефриты 1970 г. Болезни сердца Рак Инсульт Грипп, пневмония, бронхит | Количество всех смертных случаев, % 14,4 11,3 8,1 8,0 18,9 9,4 8,6 8,0 38,3 17,2 10,6 3,6 |
Относительно скромные значения средней продолжительности жизни (35—40 лет) в сравнении с максимально регистрируемой (115—120 лет) в первой третиXX в. в подавляющем большинстве стран Европы, отличавшихся в тот период относительно высоким жизненным уровнем, затрудняли прицельное изучение влияния физико-химических, климатических и иных природных факторов на скорость старения в популяциях, занимающих различные территории. В настоящее время с увеличением средней длительности жизни до 67,3—74,1 года у мужчин и до 74,2—79,9 года у женщин ситуация изменилась.
Рассмотрим следующий пример. Мужчин Финляндии отличает низкий показатель средней продолжительности жизни (69,2 года в 1980 г.) и один из самых высоких показателей смертности от сердечно-сосудистых заболеваний в Европе. При этом в самой Финляндии особенно высоких значений этот показатель достигает в Северной Карелии. При изучении минерального состава почв и грунтовых вод этого района оказалось, что в них исключительно низка концентрация микроэлемента селена. Соответственно низкой оказалась и концентрация Se в крови финнов: 0,056—0,081 мкг/мл, в сравнении с 0,206 мкг/мл, например, у жителей США. У мужчин с концентрацией Se в сыворотке крови менее 0,045 мкг/мл риск смерти от сердечно-сосудистых заболеваний выше в 2,7 раза, а от ишемической болезни сердца — в 3,6 раза. Зависимость скорости старения от поступления в организм достаточных количеств Se объясняется тем, что последний входит в состав фермента глутаминопероксидазы. Названный фермент катализирует реакции распада Н202 и органических гидропероксидов, разрушающих мембраны и другие клеточные структуры.
Следует отдавать отчет в том, что Se представляет собой лишь один из природных факторов, который, включаясь в процессы жизнедеятельности, изменяет риск смерти, влияя на скорость возрастной перестройки сердечно-сосудистой и других систем организма. Число таких факторов велико, причем большинство из них, видимо, еще не определены. С другой стороны, в Финляндии недостаток Se в диете мало сказывается на жизнеспособности женщин, которые по средней продолжительности жизни (77,6 года в 1980 г.) относятся в Европе к достаточно благополучным популяциям. Причина, по-видимому, кроется в генотипических различиях: у мужчин во всех клетках активна единственная хромосома Х материнского происхождения со всеми ее дефектами, а у женщин в разных клетках активна хромосома Х то материнского, то отцовского происхождения.
Испытания атомного и водородного оружия, развитие ядерной энергетики и технологий повышают уровень радиационного фона. Расчеты на человечество в целом показывают, что хроническое облучение населения планеты с мощностью дозы 1 Гр на поколение сокращает продолжительность жизни на 50 000 лет на каждый миллион живых новорожденных в первом облученном поколении. В настоящее время за счет естественных и созданных человеком источников радиации за поколение человечество получает менее 0,05 Гр. Воздействие на живые ткани ионизирующего облучения приводит к образованию в них свободных радикалов, которые повышают риск развития сердечно-сосудистых заболеваний и рака (рис. 8.35). Так как нарастание риска указанных заболеваний является показателем скорости старения, изменения условий жизни в сторону повышения радиационного воздействия ускоряют названный процесс.
Определенные, хотя и ограниченные представления о влиянии условий жизни на процесс старения дает изучение субпопуляций долгожителей. Напомним, что к долгожителям причисляются лица в возрасте 90 лет и более. Их количество в популяции выражается индексом долгожительства, т.е. долей долгожителей среди населения старше 60 лет: 90/60·1000, где 90 — число долгожителей; 60 — число членов популяции в возрасте 60 лет. На большей территории бывшего СССР (названия государства и административных единиц соответствуют 1970 г.; приводимые цифры основаны на данных переписи, проведенной в указанном году) индекс долгожительства не превышает 20%. В ряде территорий, однако, его значения превосходят названное в 2,5—3 раза. Для абхазов в Абхазии, азербайджанцев в Нахичевани, балкарцев в Кабардино-Балкарии, ингушей в Северной Осетии и Чечено-Ингушетии он превосходит 60%, для эвенков в Якутии — 50%.
Из приведенных сведений видно, что долгожительство характеризует определенные этнические группы, проживающие в основном в сельской местности, для которых можно предполагать достаточно высокий уровень брачной изоляции. Это наводит на мысль о генетической основе явления или же каких-то особенностях образа жизни. Два факта тем не менее свидетельствуют в пользу того, что и условия жизни могут иметь некоторое значение. Во-первых, индекс долгожительства, например, азербайджанцев, проживающих в Грузии и Азербайджане, оказывается ниже примерно на 10%, чем для проживающих в Нахичевани. Во-вторых, русское население в зонах долгожительства (Кавказ, Якутия) отличается более низкими значениями индекса в сравнении с населением, исторически традиционным территории.
§
Геронтология знает не менее 500 гипотез, объясняющих и первопричину, и механизмы старения организма. Подавляющее большинство их не выдержало проверки временем и представляет чисто исторический интерес. К ним, в частности, относятся гипотезы, связывающие старение с расходованием особого вещества клеточных ядер, страхом смерти, утратой некоторых невосполняемых веществ, получаемых организмом в момент оплодотворения, самоотравлением продуктами жизнедеятельности, токсичностью продуктов, образуемых под действием микрофлоры толстого кишечника. Гипотезы, представляющие научную ценность в наши дни, соответствуют одному из двух главных направлений.
Некоторые авторы рассматривают старение как стохастический процесс возрастного накопления «ошибок», неизбежно случающихся в ходе обычных процессов жизнедеятельности, а также повреждений биологических механизмов под действием внутренних (спонтанные мутации) или внешних (ионизирующее облучение) факторов. Стохастичность обусловливается случайным характером изменений во времени и локализации в организме. В различных вариантах гипотез данного направления первостепенная роль отводится разным внутриклеточным структурам, от первичного повреждения которых зависят функциональные расстройства на клеточном, тканевом и органном уровнях. Прежде всего это генетический аппарат клеток (гипотеза соматических мутаций).
Многие исследователи связывают начальные изменения старения организма с изменениями строения и, следовательно, физико-химических и биологических свойств макромолекул: ДНК, РНК, белков хроматина, цитоплазматических и ядерных белков, ферментов. Особо выделяют также липиды клеточных мембран, часто являющиеся мишенью для свободных радикалов. Сбои в работе рецепторов, в частности клеточных оболочек, нарушают эффективность регуляторных механизмов, что приводит к рассогласованию процессов жизнедеятельности.
К рассматриваемому направлению относятся также гипотезы, усматривающие первооснову старения в нарастающем с возрастом износе структур в диапазоне от макромолекул до организма в целом, приводящем в конце концов к состоянию, не совместимому с жизнью. Такое представление, однако, слишком прямолинейно. Напомним, что возникновению и накоплению мутационных изменений в ДНК противостоят природные антимутационные механизмы, а вредные последствия образования свободных радикалов снижаются благодаря функционированию антиоксидантных механизмов. Таким образом, если «концепция износа» биологических структур правильно отражает сущность старения, то итог в виде большей или меньшей скорости старческих изменений возраста, в котором у разных людей эти изменения становятся очевидными, является следствием наложения разрушительных и защитных процессов. В этом случае гипотеза износа с неизбежностью включает в себя такие факторы, как генетическая предрасположенность, условия и даже образ жизни, от которых, как мы видели, зависит скорость старения.
Второе направление представлено генетическими или программными гипотезами, согласно которым процесс старения находится под прямым генетическим контролем. Указанный контроль, согласно одним взглядам, осуществляется с помощью специальных генов. По другим взглядам, он связан с наличием специальных генетических программ, как это имеет место в отношении других стадий онтогенеза, например эмбриональной.
В пользу запрограммированности старения приводят доказательства, многие из которых уже рассмотрены в разд. 8.6.1. Обычно также ссылаются на наличие в природе видов, у которых вслед за размножением бурно нарастают изменения, приводящие животных к гибели. Типичный пример —тихоокеанские лососи (нерка, горбуша), погибающие после нереста. Пусковой механизм в этом случае связан с изменением режима секреции половых гормонов, что следует рассматривать как особенность генетической программы индивидуального развития лососевых, отражающей их экологию, а не как универсальный механизм старения. Примечательно, что кастрированная горбуша не нерестится и живет в 2—3 раза дольше. Именно в эти дополнительные годы жизни следует ожидать появления признаков старения в клетках и тканях.
Некоторые программные гипотезы основаны на допущении, что в организме функционируют биологические часы, в соответствии с которыми происходят возрастные изменения. Роль «часов» приписывают, в частности, вилочковой железе, прекращающей функционирование при переходе организма в зрелый возраст. Еще один кандидат — это нервная система, особенно некоторые ее отделы (гипоталамус, симпатическая нервная система), главным функциональным элементом которой являются первично стареющие нервные клетки. Допустим, что прекращение в определенном возрасте функций тимуса, что, несомненно, находится под генетическим контролем, является сигналом начала старения организма. Это, однако, не означает генетического контроля процесса старения. В отсутствие тимуса ослабляется иммунологический контроль за аутоиммунными процессами. Но для того чтобы эти процессы пошли, необходимы либо мутантные лимфоциты (повреждения ДНК), либо белки с измененной структурой и антигенными свойствами.
Генетические программы, в том числе и индивидуального развития, являются всегда результатом эволюции, закрепляемым в генофонде вида вследствие естественного отбора. На первый взгляд, естественный отбор должен благоприятствовать увеличению продолжительности жизни. В связи с этим приобретение видом в ходе эволюции генетической программы старения, обусловливающего неизбежность смерти, представляется маловероятным. Рассмотрим следующий пример. В природных условиях в первый год жизни сохраняется в живых лишь 1/4 синиц каждого поколения. По истечении 2-го года от поколения остаются единицы, если это вообще происходит. В лабораторных условиях птицы достигают 9-летнего возраста. В таком случае практически невозможно объяснить, в силу каких обстоятельств естественный отбор мог формировать генетическую программу саморазрушения организма в процессе старения, рассчитанную на 7—8 лет жизни, которые синицами не проживаются.
Изложенное выше не исключает зависимости скорости старения и времени наступления старческих изменений от генетических факторов, однако этими факторами не являются специальные гены или программа. Рассмотрим еще один пример. Для хореи Гентингтона типичным признаком служит сильный тремор (дрожание) головы, конечностей (пляска святого Витта). Симптомы этого наследственного заболевания обычно появляются в возрасте 35—39 лет, причем у мужчин позднее, чем у женщин. Различие в сроках появления болезни объясняется особенностями эволюции мужского и женского генотипов. У мужчин, имеющих по сравнению с женщинами большую продолжительность репродуктивного периода, давление отбора против соответствующего признака угасает с возрастом более медленно. Неблагоприятное фенотипическое действие гена, лежащего в основе хореи Геттингтона, в юношеском и зрелом возрасте подавлялось благодаря присутствию в геноме генов-модификаторов (см. разд. 8.6.1).
Таким образом, из двух принципиально различных направлений в объяснении старения как закономерной стадии онтогенеза в настоящее время более обоснованным является представление, рассматривающее этот процесс как износ биологических структур, а не генетически предопределенное саморазрушение.
67.Основные концепции в биологии развития (преформизм, эпигенез).
Вопрос об индивидуальном развитии — онтогенезе — привлекал к себе внимание со времен Аристотеля. Благодаря усилиям многих исследователей к XVII в. был накоплен обширный материал по изменениям, происходящим с зародышами позвоночных на макроуровне. Появление в XVII столетии [микроскопа] перевело эмбриологию на качественно новый уровень, хотя несовершенство первых микроскопов и крайне примитивная техника изготовления микропрепаратов делали практически недоступными для изучения ранние стадии развития зародышей.
В XVII-XVIII вв. оформились два взгляда на онтогенез — преформизм и эпигенез. Сторонники преформизма полагали, что зародышевое развитие сводится к росту вполне сформированного зародыша. Предполагалось, что зародыш — уменьшенный вариант сложноустроенного взрослого организма — существовал в такой форме с момента творения. Преформисты, в свою очередь, разделились на две группы. Овисты — Я. Сваммердам, А. Валлиснери, М. Мальпиги, Ш. Бонне, А. Галлер, Л. Спаланзани и др. — считали, что уже сформированный зародыш находится в яйцеклетке, а мужское половое начало лишь дает толчок к развитию. Анималькулисты (А. Левенгук, Н. Хартсекер, И.Н. Либеркюн и др.) утверждали, что зародыш заключен в сперматозоиде, который развивается за счет питательных веществ яйца. А. Левенгук допускал существование “мужских” и “женских” сперматозоидов. Крайним выражением преформизма явилась теория вложений. Согласно ей, половые клетки зародышей, как матрешки, уже несут в себе зародышей следующего поколения, в тех содержатся зародыши последующих поколений, и так далее.
Воззрения преформистов базировались на некоторых фактических данных. Так, Я. Сваммердам, вскрыв куколку бабочки, обнаружил там вполне сформированное насекомое. Ученый воспринял это как доказательство того, что более поздние стадии развития “спрятаны” в более ранних и до поры до времени не видны. Ш. Бонне открыл партеногенез — размножение без оплодотворения — у тлей, что было воспринято овистами как довод в их пользу.
Ряд фактов с точки зрения преформизма получал не слишком удачное объяснение. В XVIII в. благодаря работам Р. Реомюра и А. Трамбле становится широко известным явление регенерации. Ш. Бонне сводил и регенерацию, и различные формы бесполого размножения к “предсуществованию” в организме особых, специально на сей предмет предобразованных зародышей. Таким образом, гидра вся целиком с точки зрения преформизма представляла собой яичник или собрание зародышей. Сходство детей с обоими родителями преформисты объясняли тем, что зародыш, происшедший из яйца или из семенного анималькуля, формируется по образу и подобию своих родителей под влиянием воображения матери в течение утробной жизни. Плохо согласовывалось с идеей преформизма и появление уродств. Впрочем, некоторые сторонники этой концепции допускали, что вложенные зародыши не обязательно идентичны друг с другом, вплоть до того, что и сам прогресс живых форм мог быть преформирован в момент творения.
Приверженцы альтернативного течения — эпигенетики — считали, что в процессе онтогенеза происходит новообразование структур и органов зародыша из бесструктурного вещества. Впервые идея эпигенеза встречается в труде В. Гарвея “Исследования о нарождении животных” (1651 г.), но в полной мере соответствующие взгляды были выражены К.Ф. Вольфом (1733-1794). К.Ф. Вольф исходил из того, что, если правы преформисты, то все органы зародыша, как только мы их можем увидеть, должны быть полностью сформированы. В своей работе “Теория зарождения” (1759 г.) ученый описывает картины постепенного возникновения различных органов из “неорганизованной массы” у животных и растений. К сожалению, К.Ф. Вольф работал с довольно плохим микроскопом, что породило многие фактические неточности, но это не умаляет значения созданной им теории эпигенеза.
Эпигенетической точки зрения в XVIII в. придерживались П. Мопертюи, Дж. Нидхэм, Д. Дидро, отчасти Ж. Бюффон.
Решающий перелом в споре между представителями двух течений произошел в XIX в. после работ К.М. Бэра (1792-1876), сумевшего снять альтернативу — или преформизм, или эпигенез. К.М. Бэр считал, что нигде в зародыше не происходит новообразований, имеют место лишь преобразования. При этом преобразование К.М. Бэр понимал отнюдь не в духе преформизма, а рассматривал его как подлинное развитие, с глубокими качественными преобразованиями от более простого и недифференцированного к более сложному и дифференцированному.
68.Механизмы онтогенеза на клеточном уровне: пролиферация, миграция, адгезия, избирательная сортировка клеток, дифференцировка, запрограммированная гибель клеток).
69.Взаимодействие частей развивающегося организма. Эмбриональная индукция, её виды.
70.Целостность онтогенеза. Эмбриональная регуляция. Детерминация частей развивающегося организма. Морфогенез.
71.Роль наследственности и среды в онтогенезе. Критические периоды в онтогенезе человека. Тератогенез, канцерогенез. Аномалии и пороки развития.
72.Пороки развития в пренатальном периоде онтогенеза человека. Классификация пороков развития. Наследственные и ненаследственные пороки. Фенокопии.
73.Регенерация. Физиологическая регенерация, её значение.
Регенерация – процесс вторичного развития органа или ткани, вызванный повреждениями какого – либо рода.
Физиологическая регенерация – восстановление частей организма, износившихся в процессе жизнедеятельности. Действует на протяжении всего онтогенеза, поддерживает постоянство структур, несмотря на гибель клеток. Интенсивные процессы физиологической регенерации при восстановлении клеток крови, эпидермиса, слизистых оболочек. Примерами могут быть линька птиц, рост зубов у грызунов. Физиологическая регенерация происходит не только в тканях с интенсивно делящимися клетками, но и там, где клетки делятся незначительно. 25 гепатоцитов из 1000 погибают и столько же восстанавливаются. Физиологическая регенерация – динамический процесс, который включает в себя клеточное деление и другие процессы. Обеспечение функций лежит в основе нормального функционирования организма.
Физиологическая регенерация представляет собой процесс обновления функционирующих структур организма. Благодаря физиологической регенерации поддерживается структурный гомеостаз и обеспечивается возможность постоянного выполнения органами их функций. С общебиологической точки зрения, физиологическая регенерация, как и обмен веществ, является проявлением такого важнейшего свойства жизни, как самообновление.
Примером физиологической регенерации на внутриклеточном уровне являются процессы восстановления субклеточных структур в клетках всех тканей и органов. Значение ее особенно велико для так называемых «вечных» тканей, утративших способность к регенерации путем деления клеток. В первую очередь это относится к нервной ткани.
Примерами физиологической регенерации на клеточном и тканевом уровнях являются обновление эпидермиса кожи, роговицы глаза, эпителия слизистой кишечника, клеток периферической крови и др. Обновляются производные эпидермиса — волосы и ногти. Это так называемая пролиферативная регенерация, т.е. восполнение численности клеток за счет их деления. Во многих тканях существуют специальные камбиальные клетки и очаги их пролиферации. Это крипты в эпителии тонкой кишки, костный мозг, пролиферативные зоны в эпителии кожи. Интенсивность клеточного обновления в перечисленных тканях очень велика. Это так называемые «лабильные» ткани. Все эритроциты теплокровных животных, например, сменяются за 2—4 мес, а эпителий тонкой кишки полностью сменяется за 2 сут. Это время требуется для перемещения клетки из крипты на ворсинку, выполнения ею функции и гибели. Клетки таких органов, как печень, почка, надпочечник и др., обновляются значительно медленнее. Это так называемые «стабильные» ткани.
Об интенсивности пролиферации судят по количеству митозов, приходящихся на 1000 подсчитанных клеток. Если учесть, что сам митоз в среднем длится около 1 ч, а весь митотаческий цикл в соматических клетках в среднем протекает 22—24 ч, то становится ясно, что для определения интенсивности обновления клеточного состава тканей необходимо подсчитать количество митозов в течение одних или нескольких суток. Оказалось, что количество делящихся клеток не одинаково в разные часы суток. Так был открыт суточный ритм клеточных делений.
Суточный ритм количества митозов обнаружен не только в нормальных, но и в опухолевых тканях. Он является отражением более общей закономерности, а именно ритмичности всех функций организма. Одна из современных областей биологии — хронобиология — изучает, в частности, механизмы регуляции суточных ритмов митотической активности, что имеет весьма важное значение для медицины. Существование самой суточной периодичности количества митозов указывает на регулируемость физиологической регенерации организмом. Кроме суточных существуют лунные и годичные циклы обновления тканей и органов.
В физиологической регенерации выделяют две фазы: разрушительную и восстановительную. Полагают, что продукты распада части клеток стимулируют пролиферацию других. Большую роль в регуляции клеточного обновления играют гормоны.
Физиологическая регенерация присуща организмам всех видов, но особенно интенсивно она протекает у теплокровных позвоночных, так как у них вообще очень высока интенсивность функционирования всех органов по сравнению с другими животными.
74.Репаративная регенерация, её значение. Способы репаративной регенерации. Проявление регенерационной способности в филогенезе. Молекулярно-генетические, клеточные и системные механизмы регенерации. Особенности восстановительных процессов у млекопитающих.
Регенерация может быть физиологической и репаративной. В свою очередь репаративная регенерация бывает нескольких видов:
– возмещающая;
– посттравматическая;
– восстановительная;
– патологическая.
Репаративная регенерация – восстановление поврежденных тканей и органов после чрезвычайных воздействий. При полной регенерации восстанавливается полное исходное строение ткани после ее повреждения, её архитектура остается неизменной. Распространена у организмов, способных к бесполому размножению. Например, белая планария, гидра, моллюски (если удалить голову, но оставить нервно – узловую структуру). Типичная репаративная регенерация возможна у высших организмов, в т.ч. и человека. Например, при устранении некротических клеток органов. В острой стадии пневмонии происходит деструкция альвеол и бронхов, затем происходит восстановление. При действии гепатотропных ядов возникают диффузные некротические изменения печени. После прекращения действия ядов восстанавливается архитектоника за счет деления гепатоцитов – клеток печеночной паренхимы. Восстанавливается исходная структура. Гомоморфоз – восстановление структуры в том виде, в котором она существовала до разрушения. Неполная репаративная регенерация – регенерированный орган отличается от удаленного – гетероморфоз. Исходная структура не восстанавливается, а иногда вместо одного органа развивается другой орган. Например, глаз у рака. При удалении в некоторых случаях развивается антенна. У человека печень при удалении части печеночной доли аналогично регенерирует. Возникает рубец и через 2 – 3 месяца после операции масса печени восстанавливается, а восстановления формы органа не происходит. Это происходит из-за удаления и повреждения соединительной ткани во время операции.
У млекопитающих могут регенерировать все 4 вида ткани.
1. Соединительная ткань. Рыхлая соединительная ткань обладает высокой способностью к регенерации. Лучше всего регенерируют интерстициальные компоненты – образуется рубец, замещающийся тканью. Костная ткань – аналогично. Основные элементы, восстанавливающие ткань – остеобласты (малодифференцированные камбиальные клетки костной ткани);
2. Эпителиальная ткань. Обладает выраженной регенерационной реакцией. Эпителий кожи, роговая оболочка глаза, слизистые оболочки полости рта, губ, носа, желудочно-кишечного тракта, мочевого пузыря, слюнные железы, паренхима почек. При наличии раздражающих факторов могут происходить патологические процессы, приводящие к разрастанию тканей, что приводит к раковым опухолям.
3. Мышечная ткань. Значительно меньше регенерирует, чем эпителиальная и соединительная ткани. Поперечная мускулатура – амитоз, гладкая – митоз. Регенерирует за счет недифференцированных клеток – сателлитов. Могут разрастаться и регенерировать отдельные волокна, и даже целые мышцы.
4. Нервная ткань. Обладает плохой способностью к регенерации. В эксперименте показано, что клетки периферической и вегетативной нервной системы, двигательные и чувствительные нейроны в спинном мозге мало регенерируют. Аксоны хорошо регенерируют за счет Шванновских клеток. В головном мозге вместо них – глия, поэтому регенерация не происходит.
При регенерации миокарда и центральной нервной системы сначала образуется рубец, а затем идет регенерация за счет увеличения размеров клеток, внутриклеточная регенерация также имеет место. Клетки миокарда митозом не делятся. Разница происходит из-за развития в эмбриональном периоде. У взрослых организмов очень мощно функционирует ЭПР и это тормозит клеточное деление.
Процесс регенерации конечности у тритона/ саламандры.
После ампутации регенерация конечности происходит строго упорядоченно, всегда одинаково. Восстанавливающийся конец округляется, затем приобретает коническую форму, растет в длину, становится похожим на ласт. Потом закладываются пальцы. К 8 неделе регенерация конечности полностью завершена.
На клеточном уровне выделяют несколько фаз регенерации конечности:
1) фаза заживления раны;
2) процесс демонтирования;
3) фаза « конической бластемы»;
4) фаза редифференцировки.
Фаза заживления раны. В этот период происходит обрастание клетками раны на культе, возникает апикальная «шапочка» (если контакт нарушен – регенерации не будет).
Процесс демонтирования. После заживления, в тканях, прилежащих к культе, происходит рассасывание ткани. Мышечные волокна утрачивают упорядоченность, становятся «растрепанными». В костной ткани утрачивается надкостница, появляются гигантские фагоцитирующие клетки, имеющие не менее 3-х ядер. Эти клетки захватывают матрикс и освобождают место для роста новой кости и хряща, удаляя ненужный материал. Концевая часть культи становится отечной и выпячивается. В культе накапливаются однотипные дедифференцированные клетки, уподобленные эмбриональным клеткам. Через некоторое время начинается деление дедифференцированных клеток.
В отрастающую культю врастают нервы, и наступает стадия « конической бластемы». Конечность имеет форму ласта, нарастает клеточная масса, восстанавливается кровоток. Возникает «регенерационная почка».
Фаза редифференцировки. Конечность удлиняется, начинается редифференцировка, и процесс регенерации подходит к концу. Если денервировать конечность – регенерация не произойдет т.к. нервная ткань выполняет эндокринную, проводящую функции. Кроме того, нервная ткань осуществляет секрецию белкового гормона, под контролем которого осуществляется регенерация.
Репаративная (от лат. reparatio — восстановление) регенерация наступает после повреждения ткани или органа. Она очень разнообразна по факторам, вызывающим повреждения, по объемам повреждения, по способам восстановления. Механическая травма, например оперативное вмешательство, действие ядовитых веществ, ожоги, обморожения, лучевые воздействия, голодание, другие болезнетворные агенты,— все это повреждающие факторы. Наиболее широко изучена регенерация после механической травмы. Способность некоторых животных, таких, как гидра, планария, некоторые кольчатые черви, морские звезды, асцидия и др., восстанавливать утраченные органы и части организма издавна изумляла ученых. Ч. Дарвин, например, считал удивительными способность улитки воспроизводить голову и способность саламандры восстанавливать глаза, хвост и ноги именно в тех местах, где они отрезаны.
Объем повреждения и последующее восстановление бывают весьма различными. Крайним вариантом является восстановление целого организма из отдельной малой его части, фактически из группы соматических клеток. Среди животных такое восстановление возможно у губок и кишечнополостных. Среди растений возможно развитие целого нового растения даже из одной соматической клетки, как это получено на примере моркови и табака. Такой вид восстановительных процессов сопровождается возникновением новой морфогенетической оси организма и назван Б.П. Токиным «соматическим эмбриогенезом», ибо во многом напоминает эмбриональное развитие.
Существуют примеры восстановления больших участков организма, состоящих из комплекса органов. В качестве примера служат регенерация ротового конца у гидры, головного конца у кольчатого червя и восстановление морской звезды из одного луча (рис. 8.24). Широко распространена регенерация отдельных органов, например конечности у тритона, хвоста у ящерицы, глаз у членистоногих. Заживление кожных покровов, ран, повреждений костей и других внутренних органов является менее объемным процессом, но не менее важным для восстановления структурно-функциональной целостности организма. Особый интерес представляет способность зародышей на ранних стадиях развития восстанавливаться после значительной утраты материала. Эта способность была последним аргументом в борьбе между сторонниками преформизма и эпигенеза и привела в 1908 г. Г. Дриша к концепции эмбриональной регуляции.
Рис. 8.24. Регенерация комплекса органов у некоторых видов беспозвоночных животных. А — гидра;Б — кольчатый червь; В — морская звезда
(пояснение см. в тексте)
Существует несколько разновидностей или способов репаративной регенерации. К ним относят эпиморфоз, морфаллаксис, заживление эпителиальных ран, регенерационную гипертрофию, компенсаторную гипертрофию.
Эпителизация при заживлении ран с нарушенным эпителиальным покровом идет примерно одинаково, независимо от того, будет далее происходить регенерация органа путем эпиморфоза или нет. Эпидермальное заживление раны у млекопитающих в том случае, когда раневая поверхность высыхает с образованием корки, проходит следующим образом (рис. 8.25). Эпителий на краю раны утолщается вследствие увеличения объема клеток и расширения межклеточных пространств. Сгусток фибрина играет роль субстрата для миграции эпидермиса в глубь раны. В мигрирующих эпителиальных клетках нет митозов, однако они обладают фагоцитарной активностью. Клетки с противоположных краев вступают в контакт. Затем наступает кератинизация раневого эпидермиса и отделение корки, покрывающей рану.

Рис. 8.25. Схема некоторых событий, происходящих
при эпителизации кожной раны у млекопитающих.
А—начало врастания эпидермиса под некротическую ткань; Б— срастание эпидермиса и отделение струпа:
1—соединительная ткань, 2—эпидермис, 3—струп, 4—некротическая ткань
К моменту встречи эпидермиса противоположных краев в клетках, расположенных непосредственно вокруг края раны, наблюдается вспышка митозов, которая затем постепенно падает. По одной из версий, эта вспышка вызвана понижением концентрации ингибитора митозов — кейлона.
Эпиморфоз представляет собой наиболее очевидный способ регенерации, заключающийся в отрастании нового органа от ампутационной поверхности. Регенерация конечности тритона и аксолотля изучена детально. Выделяют регрессивную и прогрессивную фазы регенерации. Регрессивная фаза начинается с заживления раны, во время которого происходят следующие основные события: остановка кровотечения, сокращение мягких тканей культи конечности, образование над раневой поверхностью сгустка фибрина и миграция эпидермиса, покрывающего ампутационную поверхность.
Затем начинается разрушение остеоцитов на дистальном конце кости и других клеток. Одновременно в разрушенные мягкие ткани проникают клетки, участвующие в воспалительном процессе, наблюдается фагоцитоз и местный отек. Затем вместо образования плотного сплетения волокон соединительной ткани, как это происходит при заживлении ран у млекопитающих, в области под раневым эпидермисом утрачиваются дифференцированные ткани. Характерна остеокластическая эрозия кости, что является гистологическим признаком дедифференцировки. Раневой эпидермис, уже пронизанный регенерирующими нервными волокнами, начинает быстро утолщаться. Промежутки между тканями все более заполняются мезенхимоподобными клетками. Скопление мезенхимных клеток под раневым эпидермисом является главным показателем формирования регенерационной бластемы. Клетки бластемы выглядят одинаково, но именно в этот момент закладываются основные черты регенерирующей конечности.
Затем начинается прогрессивная фаза, для которой наиболее характерны процессы роста и морфогенеза. Длина и масса регенерационной бластемы быстро увеличиваются. Рост бластемы происходит на фоне идущего полным ходом формирования черт конечности, т.е. ее морфогенеза. Когда форма конечности в общих чертах уже сложилась, регенерат все еще меньше нормальной конечности. Чем крупнее животное, тем больше эта разница в размерах. Для завершения морфогенеза требуется время, по истечении которого регенерат достигает размеров нормальной конечности.
Некоторые стадии регенерации передней конечности у тритона после ампутации на уровне плеча показаны на рис. 8.26. Время, необходимое для полной регенерации конечности, варьирует в зависимости от размера и возраста животного, а также от температуры, при которой она протекает.

Рис. 8.26. Стадии регенерации передней конечности у тритона
У молодых личинок аксолотлей конечность может регенерировать за 3 нед, у взрослых тритонов и аксолотлей за 1—2 мес, а у наземных амбистом для этого требуется около 1 года.
При эпиморфной регенерации не всегда образуется точная копия удаленной структуры. Такую регенерацию называют атипичной. Существует много разновидностей атипичной регенерации. Гипоморфоз — регенерация с частичным замещением ампутированной структуры. Так, у взрослой шпорцевой лягушки возникает шиловидная структура вместо конечности. Гетероморфоз — появление иной структуры на месте утраченной. Это может проявляться в виде гомеозисной регенерации, заключающейся в появлении конечности на месте антенн или глаза у членистоногих, а также в изменении полярности структуры. Из короткого фрагмента планарии можно стабильно получать биполярную планарию (рис. 8.27).
Встречается образование дополнительных структур, или избыточная регенерация. После надреза культи при ампутации головного отдела планарии возникает регенерация двух голов или более (рис. 8.28). Можно получить больше пальцев при регенерации конечности аксолотля, повернув конец культи конечности на 180°. Дополнительные структуры являются зеркальным отражением исходных или регенерировавших структур, рядом с которыми они расположены (закон Бэйтсона).

Рис. 8.27. Биполярная планария
Морфаллаксис — это регенерация путем перестройки регенерирующего участка. Примером служит регенерация гидры из кольца, вырезанного из середины ее тела, или восстановление планарии из одной десятой или двадцатой ее части. На раневой поверхности в этом случае не происходит значительных формообразовательных процессов. Отрезанный кусочек сжимается, клетки внутри него перестраиваются, и возникает целая особь уменьшенных размеров, которая затем растет. Этот способ регенерации впервые описал Т. Морган в 1900 г. В соответствии с его описанием морфаллаксис осуществляется без митозов. Нередко имеет место сочетание эпиморфного роста на месте ампутации с реорганизацией путем морфаллаксиса в прилежащих частях тела.

Рис. 8.28. Многоголовая планария, полученная после ампутации головы
и нанесения насечек на культю
Регенерационная гипертрофия относится к внутренним органам. Этот способ регенерации заключается в увеличении размеров остатка органа без восстановления исходной формы. Иллюстрацией служит регенерация печени позвоночных, в том числе млекопитающих. При краевом ранении печени удаленная часть органа никогда не восстанавливается. Раневая поверхность заживает. В то же время внутри оставшейся части усиливается размножение клеток (гиперплазия) и в течение двух недель после удаления 2/3 печени восстанавливаются исходные масса и объем, но не форма. Внутренняя структура печени оказывается нормальной, дольки имеют типичную для них величину. Функция печени также возвращается к норме.
Компенсаторная гипертрофия заключается в изменениях в одном из органов при нарушении в другом, относящемся к той же системе органов. Примером является гипертрофия в одной из почек при удалении другой или увеличение лимфатических узлов при удалении селезенки.
Последние два способа отличаются местом регенерации, но механизмы их одинаковы: гиперплазия и гипертрофия.
Восстановление отдельных мезодермальных тканей, таких, как мышечная и скелетная, называют тканевой регенерацией. Для регенерации мышцы важно сохранение хотя бы небольших ее культей на обоих концах, а для регенерации кости необходима надкостница. Регенерация путем индукции происходит в определенных мезодермальных тканях млекопитающих в ответ на действие специфических индукторов, которые вводят внутрь поврежденной области. Этим способом удается получить полное замещение дефекта костей черепа после введения в него костных опилок.
Таким образом, существует множество различных способов или типов морфогенетических явлений при восстановлении утраченных и поврежденных частей организма. Различия между ними не всегда очевидны, и требуется более глубокое понимание этих процессов.
Изучение регенерационных явлений касается не только внешних проявлений. Существует целый ряд вопросов, носящих проблемный и теоретический характер. К ним относятся вопросы регуляции и условий, в которых протекают восстановительные процессы, вопросы происхождения клеток, участвующих в регенерации, способности к регенерации у различных групп, животных и особенностей восстановительных процессов у млекопитающих.
Установлено, что в конечности амфибий после ампутации и в процессе регенерации происходят реальные изменения электрической активности. При проведении электрического тока через ампутированную конечность у взрослых шпорцевых лягушек наблюдается усиление регенерации передних конечностей. В регенератах увеличивается количество нервной ткани, из чего делается вывод, что электрический ток стимулирует врастание нервов в края конечностей, в норме не регенерирующих.
Попытки стимулировать подобным образом регенерацию конечностей у млекопитающих оказались безуспешными. Так, под действием электрического тока или при сочетании действия электрического тока с фактором роста нервов удавалось получить у крысы только разрастание скелетной ткани в виде хрящевых и костных мозолей, которые не походили на нормальные элементы скелета конечностей.
Несомненна регуляция регенерационных процессов со стороны нервной системы. При тщательной денервации конечности во время ампутации эпиморфная регенерация полностью подавляется и бластема никогда не образуется. Были проведены интересные опыты. Если нерв конечности тритона отвести под кожу основания конечности, то образуется дополнительная конечность. Если его отвести к основанию хвоста — стимулируется образование дополнительного хвоста. Отведение нерва на боковую область никаких дополнительных структур не вызывает. Эти эксперименты привели к созданию концепции регенерационных полей. .
§
В процессе эволюции закономерным является как возникновение новых структур, так и их исчезновение. В основе лежит принцип дифференциации, проявляющийся на фоне первичной полифункциональности и способности функций изменяться количественно. Любая структура при этом возникает на основе предшествующих структур вне зависимости от того, на каком уровне организации живого осуществляется процесс филогенеза. Так, известно, что около 1 млрд. лет назад исходный белок глобин вслед за дупликацией исходного гена дифференцировался на мио- и гемоглобин — белки, входящие в состав соответственно мышечных и кровяных клеток и дифференцировавшиеся в связи с этим по функциям. В филогенезе центральной нервной системы хордовых также можно видеть дифференцировку и смену функций структур: головной мозг формируется из переднего конца нервной трубки. Таким же образом новые биологические виды образуются в виде изолированных популяций исходных видов (см. § 11.6), а новые биогеоценозы — за счет дифференцировки предсуществующих (см. § 16.2).
В связи с тем что ниже будут рассмотрены филогенезы конкретных систем органов, подробнее остановимся на закономерностях возникновения и исчезновения органов. Примером возникновения органов служит происхождение матки плацентарных млекопитающих от парных яйцеводов. При удлинении эмбрионального развития млекопитающих возникает необходимость более длительной задержки зародыша в организме матери. Это может осуществляться только в каудальных отделах яйцеводов, полость которых при этом увеличивается, а стенка дифференцируется таким образом, что к ней прикрепляется плацента, обеспечивающая взаимосвязь организма матери и плода. В процессе естественного отбора сохранялись и успешно размножались в первую очередь те млекопитающие, в организмах самок которых потомство развивалось наиболее долго. В итоге возник новый орган — матка, обеспечивающий зародышу оптимальные условия внутриутробного развития (см. разд. 14.5.3) и повышающий выживаемость соответствующих видов.
В возникновении такого более сложного и специализированного органа, как глаз, наблюдаются те же закономерности. В основе формирования органа зрения, как и всех органов чувств, лежат клетки кожного эпителия, среди которых дифференцируются и рецепторные, в частности светочувствительные. Объединение их в группы приводит к возникновению примитивных обособленных органов зрения, позволяющих животным лишь оценивать освещенность. Погружение такого светочувствительного органа под кожу обеспечивает сохранность нежных клеток, но при этом зрительная функция может осуществляться только благодаря возникновению прозрачности покровов. Чувствительность к свету примитивного органа зрения усиливается при утолщении прозрачных покровов и приобретении ими способностей преломлять свет и фокусировать его лучи на чувствительных клетках глаза. Сложный орган требует вспомогательного аппарата — защитных структур, мышц, приводящих его в движение, и т.д. Возросший уровень сложности организации глаза с необходимостью сопровождается усложнением регуляции его функций, что и выражается в усилении его интеграции как целостной системы.
Исчезновение, или редукция, органа в филогенезе может быть связана с тремя разными причинами и имеет различные механизмы. Во-первых, орган, выполнявший ранее важные функции, может оказаться в новых условиях вредным. Против него срабатывает естественный отбор, и орган довольно быстро может полностью исчезнуть. Примеров такого прямого исчезновения органов немного. Так, многие насекомые малых океанических островов бескрылы вследствие постоянной элиминации из их популяций летающих особей ветром. Чаще наблюдается исчезновение органов благодаря их субституции новыми структурами, выполняющими прежние функции с большей интенсивностью. Так исчезают, например, у пресмыкающихся и млекопитающих предпочки и первичные почки, заменяясь функционально вторичными почками. Таким же образом у рыб и земноводных происходит вытеснение хорды позвоночником.
Самый частый путь к исчезновению органов — через постепенное ослабление их функций. Такие ситуации возникают обычно при изменении условий существования. Орган, почти не выполняющий функций, выходит из-под контроля естественного отбора и проявляет обычно повышенную изменчивость. Возникающие изменения вызывают нарушение коррелятивных связей с другими частями организма. Благодаря этому такой орган зачастую становится вредным и против него начинает действовать естественный отбор.
В медицинской практике широко известно, что рудиментарные органы и у человека характеризуются широкой изменчивостью. Третьи большие коренные зубы, или «зубы мудрости», например, характеризуются не только значительной вариабельностью строения и размеров, но и разными сроками прорезывания, а также особой подверженностью кариесу. Иногда они вообще не прорезываются, а нередко, прорезавшись, в течение ближайших лет полностью разрушаются. То же касается и червеобразного отростка слепой кишки (аппендикса), который в норме может иметь длину от 2 до 20 см и быть расположенным по-разному (за брюшиной, на длинной брыжейке, позади слепой кишки и т.д.). Кроме того, воспаление аппендикса (аппендицит) встречается значительно чаще, чем воспалительные процессы в других отделах кишечника.
Процесс редукции органа противоположен по отношению к его нормальному морфогенезу. Прежде всего выпадают закладки таких частей органа, которые в норме формируются последними. При недоразвитии конечностей у человека обычно в первую очередь недоразвиваются фаланги I и V пальцев, закладывающиеся последними. У китообразных, совершенно лишенных задних конечностей благодаря ослаблению их функций в филогенезе, все же остаются закладки элементов тазового пояса, формирующиеся в процессе морфогенеза наиболее рано.
Исследования генетических основ редукции органов показали, что структурные гены, регулирующие морфогенез, не исчезают, в то время как существенным изменениям подвергаются гены, регулирующие время закладки рудиментарных органов, либо гены, ответственные за феномен индукционных взаимодействий в развивающемся зародыше. Действительно, при пересадках мезодермального материала дна ротовой полости зародыша ящерицы в ротовую полость развивающегося цыпленка возможно формирование у последнего зубов типичного строения, а пересадка кожной мезодермы ящерицы под эпидермис спины цыпленка приводит к формированию в нем типичных роговых чешуи вместо перьев.
Недоразвившиеся органы носят название рудиментарных или рудиментов. К рудиментам у человека относят, во-первых, структуры, потерявшие свои функции в постнатальном онтогенезе, но сохраняющиеся и после рождения (волосяной покров, мышцы ушной раковины, копчик, аппендикс как пищеварительный орган), и, во-вторых, органы, сохраняющиеся только в эмбриональном периоде онтогенеза (хорда, хрящевые жаберные дуги, правая дуга аорты, шейные ребра и др.).
Крайне редко редукция органов оказывается настолько полной, что от них не остается даже эмбриональных закладок. Примером такого полного исчезновения органов является редукция парных конечностей у большинства змей. Обычно же эмбриональные закладки органов, выполнявших серьезные функции даже у весьма отдаленных предков и давно утративших свое значение в постнатальном онтогенезе, продолжают развиваться у потомков в эмбриогенезе неопределенно долгое время. В первую очередь это относится к органам, связанным тесными морфогенетическими корреляциями с другими зачатками в зародышевом развитии. Так, хорда у всех позвоночных выполняет роль организационного центра, под контролем которого развиваются нервная трубка, позвоночник и дифференцируются сомиты. Нарушение развития даже участка хорды влечет за собой грубые пороки развития соответствующих структур.
Не менее тесными взаимными связями характеризуются элементы мочеполовой системы позвоночных. Так, мочеточники, связывающие нефроны предпочки с внешней средой у круглоротых, преобразуются у рыб и более высокоорганизованных классов позвоночных в два мочеполовых протока, выполняющих разные функции (см. разд. 14.5.3), а рудиментирующиеся нефроны первичной почки у пресмыкающихся и млекопитающих стимулируют развитие вторичной почки.
§
Различного рода нарушения эмбриогенеза могут привести к формированию у высокоорганизованных организмов и человека таких признаков, которые при нормальных условиях у них не встречаются, но присутствуют у более или менее отделенных предков. Такие признаки называют атавизмами. Если они снижают жизнеспособность и проявляются как морфологические аномалии, то их называют атавистическими или анцестральными (от франц. ancetre — предок) пороками развития.
По механизмам формирования различают три варианта атавизмов. Наиболее часто встречаются атавизмы, связанные с недоразвитием органов на тех этапах морфогенеза, когда они рекапитулировали предковое состояние. К примерам аномалий такого рода относятся двух- и трехкамерное сердце, гипоплазия, или недоразвитие диафрагмы, срединная расщелина твердого нёба, или «волчья пасть», и др. Атавизмы другого рода — результат нарушения редукции — персистирование (сохранение) и дальнейшее развитие эмбриональных структур, также рекапитулирующих морфологию, характерную для предков. К ним относят персистирование боталлова протока и правой дуги аорты (см. § 14.4), наличие ребер, связанных с шейным отделом позвоночника, боковые свищи шеи (см. разд. 14.2.1, 14.3.4). Третий тип атавистических пороков развития возникает в связи с нарушением перемещения органов в онтогенезе, результатом чего является их расположение в тех частях тела, где при нормальных условиях они находятся у предковых форм. У человека широко известны тазовое расположение почек (см. разд. 14.5.1), крипторхизм (неопущение яичника), высокое стояние плечевого пояса и др.
Во всех случаях атавизмов ведущими механизмами их возникновения являются не обратные мутации, приводящие к формированию предкового фенотипа, а, вероятно, мутации регуляторных генов, которые контролируют скорость морфогенеза и запуск процессов, направленных на редукцию органов. Действительно, для формирования любой структуры в многоклеточном организме необходимо слаженное функционирование десятков и даже сотен структурных генов. Одновременное возникновение адекватных друг другу мутаций целого комплекса таких генов крайне маловероятно. Однако на базе генных комплексов, унаследованных от предков, многие предковые структуры закономерно рекапитулируют в ходе эмбриогенеза, а время этих рекапитуляции и редукций контролируется значительно проще и небольшим количеством генов. Мутации таких регуляторных генов гораздо более вероятны.
Аллогенные аномалии и пороки развития
Атавистические пороки развития, объясняющиеся ходом предшествующей эволюции, относят к разряду филогенетических пороков. К этой же группе аномалий принадлежат и так называемые аллогенные аномалии. Это врожденные пороки, имеющие в своей основе генетические дефекты. Они встречаются одновременно у ряда родственных организмов и являются выражением закона гомологических рядов.
Н.И. Вавилова 1935 г. обратил внимание на то, что растения разных видов и родов, сходные по происхождению, обладают сходными рядами наследственной изменчивости, и объяснил эту закономерность сходством основных характеристик аллелофондов видов, еще недавно претерпевших дивергенцию. Эта закономерность была названа законом гомологических рядов.
Действие закона гомологических рядов распространяется на широкий круг биологических объектов. Попытки сопоставить гомологии генотипа с морфофункцйональными гомологиями не всегда успешны. Действительно, нет уверенности в том, что такое, с точки зрения сравнительной анатомии, бесспорно гомологическое образование, как хорда зародыша человека и личинки асцидии (животного, относящегося к подтипу Личиночно-хордовые Urochordata типа Хордовые), является абсолютно гомологичным и с позиций генетического контроля его морфогенеза. Однако изучение кариотипов цитогенетическим методом с использованием дифференциальной окраски хромосом и последовательности нуклеотидов в молекулах ДНК методом гибридизации (см. § 15.2) у близких и более отделенных видов привело к выводу о том, что закон гомологических рядов распространяется по крайней мере на виды в пределах рода, семейства, отряда, а по многим признакам — ив рамках класса. Так, известно, что ген, ответственный за синтез фактора VIII в системе свертывания крови, расположен в Х-хромосоме не только у человека, но и у приматов и даже у собак. Поэтому и у этих видов животных гемофилия встречается преимущественно у самцов, наследуясь так же, как и у человека.
В природе хорошо известен ген альбинизма, гомологичный у всех млекопитающих, а также и у других классов позвоночных. Среди млекопитающих распространен аутосомно-доминантный ген ахондроплазии, или хондродистрофической карликовости, нарушающий рост 70 длинных трубчатых костей. В морфогенезе мягких тканей морды млекопитающих и лица человека имеется срастание двух половин верхней губы за счет избирательной клеточной адгезии. У ряда травоядных животных, питающихся грубыми злаками или колючими ветками (грызуны, зайцы, верблюды), срастание верхней губы нерационально. В процессе естественного отбора у них закрепились адаптивные мутации, нарушающие клеточную адгезию верхней губы. У человека такое нарушение называют заячьей губой.
У людей аллогенные аномалии встречаются очень часто. Даже такой генетический дефект, как синдром Дауна, является аллогенной аномалией: известны случаи рождения детенышей гориллы с трисомией хромосомы, соответствующей 21-й паре хромосом человека. Симптоматика такой трисомии соответствует клинике синдрома Дауна у человека.
Точная диагностика и изучение механизмов возникновения аллогенных состояний представляют большой интерес и практически важны потому, что дают возможность использования животных с соответствующими признаками в качестве экспериментальных моделей для изучения клиники и лечения их у человека.
Организм как целое в историческом
И индивидуальном развитии.
Соотносительные преобразования органов
Рассмотренные выше филогенетические преобразования отдельных биологических структур протекают в эволюционирующих группах организмов как целостных системах. Устойчивые взаимозависимости органов и систем, проявляющиеся в филогенезе, называют координациями.
Механизмы соотносительных преобразований биологических структур различны, в связи с чем выделяют три их группы: биологические, динамические и топографические.
Биологические координации наблюдаются между структурами, непосредственно не связанными ни по функциям, ни по месту положения. Основным связующим звеном между ними являются адаптации к определенным условиям обитания. Так, у большинства эндопаразитов (см. § 18.6) сильно развиты половая система и органы прикрепления к телу хозяина, но при этом недоразвиты органы чувств и опорно-двигательный аппарат. Млекопитающие, обитающие на деревьях, обычно имеют стереоскопическое зрение и сильно развитый мозжечок. Позвоночные, которые дышат кислородом, растворенным в воде, имеют хвостовой плавник, жабры, слабо развитый передний мозг и содержат гемоглобин, способный связываться с кислородом при низком его парциальном давлении в среде. Все позвоночные, имеющие матку, обладают также совершенной системой терморегуляции.
Динамические координации выражаются во взаимном соответствии структур, связанных функционально. Тесные динамические координации имеются, например, между органами кровеносной и дыхательной систем. Так, животные, дышащие легкими, имеют трех- или четырехкамерное сердце и два круга кровообращения. Степень развитости нервных центров всегда соответствует интенсивности функционирования иннервируемых органов. Например, строение органа обоняния и обонятельные доли мозга у млекопитающих высоко развиты, в то время как у птиц примитивное строение периферической части обонятельного анализатора соответствует малым размерам обонятельных долей мозга. Это связано с тем, что в ориентации млекопитающих обоняние играет первостепенную роль, а для птиц оно не имеет большого значения.
Топографические координации проявляются между структурами, связанными друг с другом пространственно. Так, для каждого типа животного царства характерен своеобразный общий план строения, выражающийся в определенном взаимном расположении основных органов и систем. Например, у всех представителей типа Хордовые на спинной стороне тела расположена нервная трубка, под ней лежат хорда, пищеварительная трубка и брюшной кровеносный сосуд, а по бокам тела — производные мезодермы (см. разд. 13.5.4).
Все типы координации характеризуются высокой степенью устойчивости. Так, хордовые животные, известные с конца протерозойской эры, сохранили неизменными общий план строения до настоящего времени, на протяжении более 500 млн. лет. Феномен паразитизма возник значительно раньше появления хордовых, и поэтому комплекс адаптации к паразитическому образу жизни (см. § 18.6) является еще более древним. Длительно существуют и другие, более частные координации, возникающие вместе с появлением новых таксономических или экологических групп организмов.
Высокая устойчивость филогенетических координации обеспечивается целостностью онтогенеза каждой конкретной особи, развитие всех биологических структур которой протекает в строгом взаимном соответствии. Такое соответствие структур развивающегося организма в онтогенезе называют онтогенетическими корреляциями. Различают геномные, эргонтические и морфогенетические корреляции.
Геномные корреляции обеспечиваются целостностью генетической конституции развивающегося организма. Ведущими механизмами геномных корреляций являются генный баланс генотипа, сцепленное наследование генов, различные формы взаимодействия генов, а также плейотропность. Так, генные системы, регулирующие процессы пролиферации и избирательной гибели клеток на различных этапах органогенеза (см. разд, 8.2.1, 8.2.4), приводят к аллометрическому росту органов (см. разд. 8.3.3), благодаря чему появляются, например, удлиненный клюв, шея и задние конечности у большинства болотных птиц, длинная шея и ноги у жирафа, а также отличающиеся друг от друга пропорции тела у мужчин и женщин.
Морфогенетические корреляции возникают между органами, пространственно связанными между собой. Они основаны либо на феномене эмбриональной индукции (см. разд. 8.2.6), либо на общности эмбриональных закладок органов. Эти корреляции проявляются уже на ранних стадиях онтогенеза, когда еще отсутствуют функциональные связи между формирующимися органами. Так, зачаток хорды обусловливает развитие нервной трубки на спинной стороне зародыша и дифференцировку скелетогенной ткани внутренних частей сомита — склеротома в хрящ или кость, а глазной бокал (вырост переднего мозга) — формирование хрусталика при морфогенезе глаза.
Примером развития ряда структур из одного общего зачатка является формирование у млекопитающих и человека из закладок 1-й и 2-й пар жаберных дуг первичных челюстей, подъязычной кости, части хрящей гортани, шиловидного отростка черепа и трех слуховых косточек. При нарушении развития закладок жаберных дуг обычно образуется комплекс аномалий указанных структур, называемый аномаладом первых жаберных дуг.
Эргонтические корреляции обусловлены функциональной взаимозависимостью органов и частей организма. Они возникают на более поздних стадиях развития, когда органы начинают функционировать, и проявляются, например, в соответствии между степенью развитости мышцы, костных выступов, к которым она прикрепляется, и интенсивностью ее кровоснабжения. К такого рода корреляциям относят также соответствие вторичных половых признаков развитию гонад.
Ведущие корреляции в онтогенезе — геномные. В конечном счете именно они лежат в основе других корреляций, значение которых на протяжении онтогенеза меняется. Это связано с первичностью изменений генотипа в процессе филогенеза.
Система корреляций и сама подвергается эволюционным преобразованиям. В процессе эволюции видоизменяются вначале более частные корреляции, в то время как наиболее общие могут воспроизводиться в конкретных онтогенезах очень длительное время. В результате в ходе исторического развития происходит как отбор наиболее общих корреляций, имеющих значение при любых перестройках организма и в различных условиях обитания (общий план строения, соответствие между степенью развитости нервного центра и иннервируемыми органами), так и накопление локальных корреляций частного значения, формирующихся у организмов разных видов и отражающих специфику их образа жизни.
Корреляции общего значения обусловливают преемственность формообразовательных процессов в череде поколений организмов данного типа организации, а частные корреляции — многообразие конкретных форм жизни.
Между филогенетическими координациями и онтогенетическими корреляциями имеется теснейшая связь. Очевидно, что корреляции существуют и воспроизводятся в поколениях благодаря тому, что на протяжении предшествующей эволюции органов они преобразовывались скоординированно. С другой стороны, филогенетические координации в последующей эволюции организмов будут реализовываться благодаря воспроизведению онтогенетических корреляций в ходе индивидуального развития конкретных особей. Таким образом, в виде соотношения корреляций и координации проявляется диалектическое единство онто- и филогенеза как целостного процесса исторического развития живого.
Сопоставление конкретных форм соотносительных преобразований органов приводит к выводу о том, что морфогенетические корреляции и топографические координации точно так же, как эргонтические корреляции и динамические координации, взаимно обусловливают друг друга. В паре геномные корреляции — биологические координации также обнаруживается соответствие.
Осознание целостности и взаимообусловленности индивидуального и исторического развития необходимо врачу в связи с тем, что соотносительное преобразование биологических структур лежит в основе нормального развития организма человека, а нарушение даже одного незначительного компонента развивающейся системы может повлечь за собой возникновение целого комплекса нарушений в других ее элементах. С этим связано то, что практически все врожденные пороки развития, а также многочисленные генные болезни характеризуются не отдельными патологическими признаками, а синдромами, представляющими собой комплексы симптомов, связанных между собой процессом формирования.
Сердце у всех позвоночных закладывается на ранних этапах развития кпереди от глотки под челюстной дугой. В его морфогенезе участвует глотка как эмбриональный индуктор. Если это свойство глотки нарушено, тсгсердце может задержаться на двух- и трехкамерном уровне развития, при этом может быть нарушено и его перемещение в загрудинную область — шейная эктопия сердца (см. § 14.4). Эти явления — результат нарушений морфогенетических корреляций в развитии шейной области. Часто этот порок развития сопровождается нарушением отходящих от сердца сосудов (персистирование общего эмбрионального ствола, двух дуг аорты и т.д.) и недоразвитостью легких.
В возникновении данных аномалий ведущим механизмом выступает нарушение эргонтических корреляций сердце — сосуды — легкие. Первичным нарушением в описанном комплексе признаков является, вероятно, нарушение генетического контроля эмбриональной индукции, описанное ранее (см. разд. 8.2.6). Таким образом, приведенный пример иллюстрирует взаимоотношение разных форм соотносительных преобразований органов при формировании сложного комплекса патологических признаков, имеющих в целом атавистическую природу.
Примером нарушения чисто геномных корреляций является синдром Дауна. Увеличение доз генов 21-й хромосомы или ее части приводит к формированию тяжелой умственной отсталости, ослаблению тонуса мышц, аномалиям мозгового черепа и мягких частей лица, светлой пигментации волос и глаз.
Кроме филогенетических координации, подкрепляемых в каждом поколении онтогенетическими корреляциями, целостность развивающегося организма отражают и такие соотносительные преобразования органов, как субституция и гетеробатмия.
Субституция — это такое эволюционное преобразование, при котором один орган замещается другим, выполняющим обычно ту же функцию с большей интенсивностью. При этом наблюдается развитие этих органов в разных направлениях. Один обычно подвергается редукции, другой — эволюционирует прогрессивно. Так, хорда замещается позвоночником и превращается в рудиментарное образование, а первичные хрящевые челюсти позвоночных заменяются вторичными костными (см. разд. 14.2.1). Это примеры гомотопной субституции, когда новый орган возникает на месте старого. При гетеротопной субституции заменяющий орган находится на новом месте. Так, функцию печени как органа кроветворения берет на себя красный костный мозг. Выделительная функция выполняется у рыб и земноводных туловищной почкой, а у пресмыкающихся и млекопитающих — тазовой.
Гетеробатмия — это такое эволюционное преобразование, при котором в одной группе организмов обнаруживается разный уровень эволюционной продвинутое™ и специализации разных частей одного и того органа, разных органов одной и той же системы или разных частей организма. Примером может являться человек, головной мозг которого за короткое время антропогенеза претерпел колоссальные морфофизиологические изменения, в то время как пищеварительная система соответствует уровню развития других приматов.
Гетеробатмия, наблюдающаяся внутри одной и той же системы органов в разных филогенетических группах, обусловливает феномен компенсации функций, благодаря которому одни и те же экологические задачи решаются разными способами. Так, грызуны и копытные млекопитающие питаются одинаковой растительной пищей, но у первых наиболее выраженные адаптации к растительноядности проявляются в строении зубов и морфофизиологии слюнных желез, в то время как вторые на фоне примитивной зубной системы имеют высокоспециализированные желудок и кишечник. Явления гетеробатмии и, следовательно, компенсации функций имеют огромное эволюционное значение в связи с тем, что в организме, даже вступившем на путь узкой специализации, всегда остаются органы и системы относительно мало специализированные, которые при меняющихся условиях могут еще прогрессивно развиваться, раскрывая перед такими филогенетическими группами новые адаптивные возможности.
96.Филогенез наружных покровов хордовых животных. Онтофилогенетические пороки развития наружных покровов у человека.
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. разд. 23.1).

Рис. 14.1. Закладка плакоидной чешуи:
1—клетки-эмалеобразователи, 2—эпидермис, 3—эмаль, 4—склеробласты—дентинообразователи, 5—дентин, 6—сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других—сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).

Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А — зародыш в возрасте 5 недель (видны млечные линии); Б — дифференцировка пяти пар сосков; В — зародыш в возрасте 7 недель

Рис. 14.5. Атавистические аномалии развития кожи.
А — гипертрихоз; Б — полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
97.Филогенез пищеварительной системы хордовых. Онтофилогенетические пороки пищеварительной системы у человека.
Уникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.

Рис. 14.18. Кишечная трубка позвоночных (А) и ее дифференцировка в эмбриогенезе человека (Б):
а—жаберные щели в глотке; 1—ротовая полость, 2—1—5-й глоточные карманы, 3—глотка, 4—пищевод, 5—желудок, 6—двенадцатиперстная кишка, 7—дорсальная поджелудочная железа, 8—вентральная поджелудочная железа, 9— тонкая кишка. 10—клоака, 11—мочевой пузырь, 12—желточный мешок, 13—желчный пузырь, 14—печень, 15—легкие, 16—закладка щитовидной железы
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными (см. § 14.1). Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum (рис. 14.18).
Ротовая полость
Рассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.19, А— В).
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть (рис. 14.19, Г). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.

Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб (см. § 14.1). У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости (рис. 14.20). Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом.

Рис. 14.20. Переход от плакоидной чешуи к зубам по краю ротового отверстия акулы: 1— плакоидная чешуя, 2—закладка новых зубов, 3—эмаль, 4—дентин, 5—зубы
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом.
У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе (рис. 14. 21). Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.

Рис. 14.21. Сверхкомплектные зубы, прорезавшиеся у человека (указаны стрелками)
§
У ланцетника кровеносная система наиболее проста (рис. 14.28, А).Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями. На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд — печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, — в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника (рис. 14.28, Б).
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд — артериальный конус, разветвляющийся последовательно на три пары сосудов:
кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, — передними полыми венами (рис. 14.28, В).
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой — легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных (рис. 14.28, Г).
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены — рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими (рис. 14.28, Д).


Рис 14.28. Строение кровеносной системы и сердца хордовых животных.
А—ланцетника; Б—рыбы; В—земноводного; Г—пресмыкающегося; Д—млекопитающего:
1—брюшная аорта, 2—жаберные артерии, 3—сонная артерия, 4—передняя кардинальная вена, 5—спинная аорта, 6—задняя кардинальная вена, 7— печеночная вена, 8—чревная артерия, 9—воротная вена печени, 10— кювьеровы протоки, 11—сердце, 12—артериальный конус, 13—предсердие, 14—венозный синус, 15—желудочек, 16—яремная вена, 17—левая дуга аорты, 18—подключичная артерия, 19—передняя полая вена, 20—легочная артерия, 21—легочная вена, 22—задняя полая вена, 23—правая дуга аорты, 24— межжелудочковая перегородка
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным (рис. 14.29). Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.

Рис. 14.29. Развитие перегородок сердца у зародыша человека в возрасте 30 сут (А) 35 сут (Б) и 2 мес (Д): 1—межпредсердная, 2—межжелудочковая
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком (см. рис. 14.31, В). Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфо-генезе сердца большое значение имеют онтогенетические корреляции (см. § 13.4). Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
§
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных (рис. 14.30). В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.

Рис. 14.30. Эволюция артериальных жаберных дуг позвоночных. А—закладка в эмбриогенезе; Б—жаберные дуги рыб; В—жаберные дуги хвостатых земноводных- Г—жаберные дуги пресмыкающихся; Д—жаберные дуги млекопитающих:
I—VI – жаберные дуги; 1—сонные артерии, 2—боталлов проток, 3—артериальные дуги большого круга кровообращения, 4—легочные артерии
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального, или боталлова, протока (рис. 14.31, А)представляющего собой часть корня спинной аорты между 4-й и’6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке (рис. 14.31, Б). Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов — отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
Рис. 14.31. Врожденные пороки развития сердца и сосудов у человека:
А — персистирование артериального (боталлова) протока у человека:
1—легочный ствол, 2—левая дуга аорты, 3—артериальный (боталлов) проток;
Б—онтофилогенетические основы формирования пороков развития сердца и отходящих от него сосудов у человека: I—сердце зародыша человека в возрасте 5 недель; II—то же, в разрезе III—разделение артериального конуса у зародыша человека в возрасте 6 недель, IV—сердце рыбы, V—то же, в разрезе, VI—персистирование неразделенного артериального конуса у человека: 1—артериальные жаберные дуги, 2—артериальный конус, 3—предсердие, 4—кардинальные вены, 5—желудочек, 6—общая легочная артерия, 7—левая дуга аорты;
В — аномалии межпредсердной (I, II) и межжелудочковой (III)перегородок
100.Филогенез выделительной и половой систем позвоночных. Связь выделительной и половой систем у позвоночных. Эволюция почки. Эволюция половых желез. Эволюция мочеполовых протоков. Онтофилогенетические пороки развития почек, половых желез и мочеполовых протоков у человека.
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом. В эмбриогенезе закладка мочеполовой системы — нефрогонотом — формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целоми-ческих животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых (рис. 14.32).

Рис. 14.32. Взаимоотношения целома (1), половой железы (2) и общего выделительного канала (3)
Эволюция почки
Органами выделения позвоночных являются почки — парные компактные органы, структурная единица которых представлена нефроном. В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос; первичная почка—туловищная, или мезонефрос, и вторичная почка – тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки (рис. 14.33, А).
Рис. 14.33. Эволюция тефрона. А—предпочка; Б, В—первичная почка; Г—вторичная почка:
1—собирательная трубочка, 2—выделительный канадец, 3—нефростом, 4—целом, 5—капиллярный клубочек, 6—капсула, 7, 8—извитой канадец, 9—петля нефрона
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её (рис. 14.33, Б, В).
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле (рис. 14.33, Г).
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки (рис. 14.34).
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития (рис. 14.35).

Рис. 14.34. Пятинедельный зародыш человека с тремя поколениями почек:
1—предпочка, 2—первичная почка, 3—вторичная почка
Рис. 14.35. Онтофилогенетически обусловленные пороки развития почек:
1—удвоение почки, 2—удвоенный мочеточник, 3—тазовая эктопия почки, 4—надпочечники
Эволюция половых желез
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов (см. разд. 6.1.2) становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
§
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал. При развитии первичной почки этот канал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них — вольфов — вступает в связь с нефронами первичной почки. Другой — мюллеров — срастается передним концом с одним из нефронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним — впадающий в клоаку.
Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров — яйцевода. У самцов мюллеров канал редуцируется и обе функции — половую и выделительную — выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой.
У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудальная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров — становится яйцеводом (рис. 14.36). У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов и человека.
Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы. У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копулятивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.

Рис. 14.36. Эволюция почки и мочеполовых каналов.
А — нейтральное зародышевое состояние; Б — анамнии; В — амниоты:
I—самки, II—самцы; 1—предпочка, 2—первичная почка, 3—канал предпочки, 4—половая железа, 5—мюллеров канал, 6—вольфов канал, 7—мочевой пузырь, 8—клоака, 9—вторичная почка, 10—матка, 11—мочеполовой синус, 12—задняя кишка, 13—половой член, 14—мочеточник вторичной почки, 15—мужская «маточка»
В эмбриогенезе человека закладываются парные вольфовы и мюллеровы каналы. Позже в зависимости от пола происходит их редукция. Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой — utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника — эпидидимис. У плодов женского пола возможно нарушение редукции вольфовых каналов, которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки (1 случай на 1000 перинатальных вскрытии). Они развиваются как результат нарушения срастания мюллеровых каналов. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение (рис. 14.37).

Рис. 14.37. Аномалии матки и полового члена у человека. А—Д—различные формы удвоения матки и влагалища; Е—удвоение полового члена
101.Филогенез нервной системы позвоночных. Этапы эволюции головного мозга позвоночных. Онтофилогенетические пороки нервной системы у человека.
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе. Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.38). Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.

Рис 14.38. Нервная трубка ланцетника: 1 — невроцель, 2 — глазки Гессе
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок (рис. 14.39).

Рис. 14.39. Эволюция головного мозга позвоночных. А—рыба; Б—земноводное; В—пресмыкающееся; Г—млекопитающее:
1—обонятельные доли, 2—передний мозг, 3—средний мозг, 4—мозжечок, 5—продолговатый мозг, 6—промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.

Рис. 14.40. Мозговые пузыри в эмбриогенезе мозга человека. А — стадия трех мозговых пузырей; Б — стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,— отсутствие замыкания нервной трубки (рис. 14.41). Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
Рис. 14.41. Рахисхиз (платиневрия)
102.Филогенез эндокринной системы. Гормоны. Эволюционные преобразования желез внутренней секреции у хордовых животных. Онтофилогенетические пороки эндокринной системы у человека.
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами — биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
Гормоны
Химическая структура гормонов многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13).
Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус — нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые.
Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).

Рис. 14.42. Развитие гипофиза у человека. А—сагиттальный срез 4-недельного зародыша; Б—объединение зачатков гипофиза из стенки глотки и основания мозга; В—формирование трех долей гипофиза на 8-й неделе развития; Г—сформированный гипофиз:
1—крыша ротовой полости, 2—основание мозга, 3—выпячивание основания мозга (задняя доля гипофиза), 4—карман Ратке (промежуточная и передняя доли гипофиза), 5—аномалия развития (добавочные дольки передней доли в стенке глотки и внутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
У хрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабатывают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин.
В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена.
Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя — пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза.
У человека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30—40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5—6 мм и шириной 0,5—1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту 139 аномалию называют эктопией аденогипофиза, она не сопровождается патологическими проявлениями. Более опасно сохранение полости в области кармана Ратке — киста кармана Ратке. Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Больные нуждаются в оперативном вмешательстве.
Щитовидная железа, гормон которой — тироксин — регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих).
У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток в виде тяжа, полого внутри, называющегося щитоязычным протоком — canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка.
Свидетельством эмбрионального перемещения железы является и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в ее ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу.
Персистирование участков подъязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи, которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда, кисты загнаиваются и прорываются с образованием срединных свищей шеи. Не менее известны такие пороки развития, как эктопия щитовидной железы, которая может развиться в результате нарушения клеточной миграции (рис. 14.43).
Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения с щитовидной железой. Это паращитовидные железы и ультимобранхиальные тельца. Первые, выделяя гормон паратиреоидин, повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных — лишь после метаморфоза. Клетки вторых выделяют кальцитонин, являющийся антагонистом паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца являются регуляторами кальциевого обмена. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду позвоночных они постепенно перемещаются из глоточной области в сторону щитовидной железы, а у млекопитающих даже срастаются с ней. Паращитовидные железы еще сохраняют самостоятельность, а клетки ультимобранхиальных телец мигрируют между фолликулами щитовидной железы и сохраняются под названием парафолликулярных клеток. Таким образом, на примере щитовидной, паращитовидных желез и ультимобранхиальных телец видна интеграция железистых структур в сложное надерганное образование, выполняющее целый комплекс взаимосвязанных функций.

Рис. 14.43. Аномалии щитовидной железы:
1—нормальное расположение. железы, 2—место эмбриональной закладки железы, 3—варианты аномального расположения железы; стрелками показано направление перемещения зачатка щитовидной железы в эмбриогенезе
У человека известны аномалии расположения паращитовидных желез, связанные с нарушением их гетеротопий.
Надпочечники позвоночных имеют двойственное происхождение. У рыб и земноводных ткани, соответствующие мозговому и корковому веществам этих желез, расположены отдельно друг от друга. Зачатки мозгового вещества связаны по происхождению с симпатическими нервными узлами и расположены метамерно. Зачатки коркового вещества развиваются из утолщений эпителия брюшины. У наземных позвоночных мозговое и корковое вещества объединяются в компактные эндокринные железы, имеющие сложное гистологическое строение. У млекопитающих они прилежат к переднему концу почек. Мозговое вещество выделяет в основном адреналин — регулятор кровообращения и энергетического обмена, а корковое вещество — разнообразные стероидные гормоны, влияющие на минеральный, углеводный обмены и функции почек.
Эндокринная система.
В любом организме вырабатываются соединения, разносящиеся по всему организму, имеющие интегративную роль. У растений есть фитогормоны, контролирующие рост, развитие плодов, цветов, развитие пазушных почек, деление камбия и др. Фитогормоны есть у одноклеточных водорослей.
Гормоны появились у многоклеточных организмов, когда возникли специальные эндокринные клетки. Однако химические соединения, играющие роль гормонов, были и раньше. Тироксин, трийодтиронин (щитовидная железа) обнаружены у цианобактерий. Гормональная регуляция у насекомых изучена плохо.
В 1965 году Вильсон выделил инсулин из морской звезды.
Оказалось, что дать определение гормону очень трудно.
Гормон – это специфическое химическое вещество, выделяемое особыми клетками в определенном участке тела, которое поступает в кровь и затем оказывает специфическое действие на определенные клетки или органы-мишени, расположенные в других областях тела, что приводит к координации функций всего организма в целом.
Известно большое количество гормонов млекопитающих. Они делятся на 3 основные группы.
Феромоны. Выделяются во внешнюю среду. С их помощью животные принимают и передают информацию. У человека запах 14 – окситететрадекановой кислоты четко различают только женщины, достигшие половой зрелости.
Наиболее просто организованные многоклеточные организмы – например, губки тоже имеют подобие эндокринной системы. Губки состоят из 2 слоев – энтодермой и экзодермой, между ними располагается мезенхима, в которой содержатся макромолекулярные соединения, характерные для соединительной ткани более высокоорганизованных организмов. В мезенхиме есть мигрирующие клетки, некоторые клетки способны секретировать серотонии, ацетилхолин. Нервная система у губок отсутствует. Вещества, синтезируемые в мезенхиме, служат для связи отдельных частей организма. Координация осуществляется за счет перемещения клеток по мезенхиме. Есть также и перенос веществ между клетками. Заложена основа химической сигнализации, которая характерна для остальных животных. Самостоятельных эндокринных клеток нет.
У кишечнополостных имеется примитивная нервная система. Первоначально нервные клетки выполняли нейросекреторную функцию. Трофическую функцию, осуществляли контроль роста, развития организма. Затем нервные клетки стали вытягиваться и образовали длинные отростки. Секрет выделялся около органа-мишени, без переноса (т.к. не было крови). Эндокринный механизм возник раньше проводникового. Нервные клетки были эндокринными, а потом получили и проводниковые свойства. Нейросекреторные клетки был первыми секреторными клетками.
Первичноротые и вторичноротые вырабатывают одинаковые стероидные и пептидные гормоны. Принято считать, что в процессе эволюции из одних полипептидных гормонов могут возникнуть новые (мутации, дупликации генов). Дупликации менее подавляются естественным отбором, чем мутации. Многие гормоны могу синтезироваться не в одной железе, а в нескольких. Например, инсулин вырабатывается в поджелудочной железе, подчелюстной железе, 12-перстной кишке и других органах. Существует зависимость генов, контролирующих синтез гормонов от положения.
103.Филогенез иммунной систем.Эволюция живого мира состояла в образовании таких форм жизни, которые активно взаимодействуют с другими живыми организмами. Биологические организмы существуют в биотическом окружении. Они взаимодействуют с другими организмами, в биоценозе существует круговорот.
Абиотические факторы менее сильно воздействуют на организм. Эволюция живой материи привела к возникновению динамической системы взаимозависимых организмов, не могут существовать без биотического взаимодействия. Такие взаимодействия проявляются в питании и обмене веществ. Существует тенденция к усложнению связей. Современные многоклеточные организмы во многом утратили возможность синтезировать даже простые органические вещества, но научились добывать их в готовом виде. Это консументы (в том числе человек). Жизнь основана на питании продуктами биосинтеза, создание которых осуществляется продуцентами (зеленые растения). Жизнь консументов напрямую зависит от биологической продуктивности растений и от развитости растениеводства. Растениеводство- источник пищи и сырья для промышленности (в том числе фармацевтической промышленности). 40% лекарственных средств в мировой фармацевтической промышленности растительного происхождения, снижение продуктивности растений приводит к голоданию, недоеданию, ухудшению качества жизни, снижению производства и уровня общественного здоровья. Важнейший фактор, понижающий урожайность растений – микроорганизмы, грибы, бактерии. Они приспособлены к паразитизму на разных растениях, животных и человеке. Пандемия гриппа – «испанки» унесла жизни 20 млн. человек. Жертвы микробного паразитизма – все живые организмы, в том числе и сами микробы.
Почему земле до сих пор не населена одними микроорганизмами?
Почему жестокий антагонизм между живыми существами не привел к гибели живой материи еще на заре эволюции?
Почему эволюция не остановилась?
Ответы на эти и многие другие вопросы дает иммунология.
Иммунитет – невосприимчивость, устойчивость, резистентность, толерантность – способность организма противостоять агрессии со стороны других биологических видов.
1902 г. Мечников и Зильбер основали учение об иммунитете.
«Иммунитет – это совокупность всех наследственных полученных и индивидуально приобретенных организмом свойств, которые препятствуют проникновению и размножению микроорганизмов, вирусов и других патогенных объектов и действию выделяемых ими продуктов».
Наследственный иммунитет – свойства невосприимчивости, полученные от предков. Он определяется генотипом.
Индивидуальный иммунитет – приобретенный иммунитет, который вырабатывается на протяжении жизни организма.
Выделяют следующие группы факторов иммунитета:
– фагоцитарные;
– конституциональные;
– лимфоидные.
Лимфоидные факторы – наследственная способность организмов создавать иммунитет.
Конституциональные – имеют общебиологическое значение. Они присутствуют у всех организмов независимо от таксономического положения.
У беспозвоночных и позвоночных существует система фагоцитоза. У растений и простейших подобная система отсутствует. Растения защищены только наследственными свойствами. Грибы и вирусы тоже не защищены. Только у позвоночных существует приобретенная невосприимчивость благодаря наличию лимфоидной системы. При этом защитные функции лимфатической системы осуществляются специфической активностью антител и иммуноглобулинов.
Антигены – любые вещества, удовлетворяющие требованиям:
§
Термин «биосфера» введен австрийским геологом Э. Зюссом в 1875 г. для обозначения особой оболочки Земли, образованной совокупностью живых организмов, что соответствует биологической концепции биосферы. В указанном смысле названный термин используют ряд исследователей и в настоящее время.
Представление о широком влиянии живых существ на протекающие в природе процессы было сформулировано В.В. Докучаевым, который показал зависимость процесса почвообразования не только от климата, но и от совокупного влияния растительных и животных организмов.
В. И Вернадский развил это направление и разработал учение о биосфере как глобальной системе нашей планеты, в которой основной ход геохимических и энергетических превращений определяется живым веществом. Он распространил понятие биосферы не только на сами организмы, но и на среду их обитания, чем придал концепции биосферы биогеохимический смысл. Большинство явлений, меняющих в масштабе геологического времени облик Земли, рассматривали ранее как чисто физические, химические или физико-химические (размыв, растворение, осаждение, выветривание пород и т. д.). В.И. Вернадский создал учение о геологической роли живых организмов и показал, что деятельность последних представляет собой важнейший фактор преобразования минеральных оболочек планеты.
С именем В.И. Вернадского связано также формирование социально-экономической концепции биосферы, отражающей ее превращение на определенном этапе эволюции в ноосферу (см. гл. 25) вследствие деятельности человека, которая приобретает роль самостоятельной геологической силы. Учитывая системный принцип организации биосферы, а также то, что в основе ее функционирования лежат круговороты веществ и потоки энергии, современной наукой сформулированы биохимическая, термодинамическая, биогеоценотическая, кибернетическая концепции биосферы.
Биосферой называют оболочку Земли, которая населена и активно преобразуется живыми существами. Согласно В.И. Вернадскому, биосфера — это такая оболочка, в которой существует или существовала в прошлом жизнь и которая подвергалась или подвергается воздействию живых организмов. Она включает: 1) живое вещество, образованное совокупностью организмов; 2) биогенное вещество, которое создается и перерабатывается в процессе жизнедеятельности организмов (газы атмосферы, каменный уголь, нефть, сланцы, известняки и др.); 3) косное вещество, которое образуется без участия живых организмов (продукты тектонической деятельности, метеориты); 4) биокосное вещество, представляющее собой совместный результат жизнедеятельности организмов и абиогенных процессов (почвы).
Учение о биосфере создано Вернадским, опубликовано в 1926 году в труде «Биосфера».
Биолдогическая концепция.
Самые ранние представлениия о биосфере как совокупности живых организмов. Первым термин употребил Ламарк 9биосфера – область жизни).
Милль и Зюсс: « Биосфера – совокупность живых существ»
Ошибка концепции: исследователи отделяли живое от неживого.
Биогеохимическая концепция.
Немецкий ученый Малишотт выдинул идею го том, что биосфера выражается прежде всего в круговороте веществ с участием живых существ.
Докучаев писал о совместном действии климата, живых организмов, минеральной среды в образовании почыв. Наиболее распростарненная концепция.
Термодинамическая концепция.
Биосфера – система, которая обменивается с космосом энергией и подчиняется Ворому Закону термодинамики.
Кибернетическая концепция.
Биосфера – система саморегулирующаяся, которую можно моделировать и изучать протекающие в ней процессы.
Геохимическая концепция.
Живые организмы взаимодействуют между собой и с окружающей средой. Пр этом создаются глубокие метаболические связи с геохимическими факторами. Минеральные вещества идут на построении е скелета, регуляцию осмоса, функции кровеносной системы, велючается в обмен веществ. Изменяется среда – изменяется состав организмов. Дальнейшая разработка концепции принадлежит Вернадскому.
1. Его заслуга в том, что он разработал учение о биосфере как глобальной системе Земли, в которой основной ход геохимических и энергетиеских превращений определяетсяживыми организмами.
2. Распространил поняие «биосфера» на живые организмы и среду их обитания.
3. Создал теорию о живом веществе и геохимической роли живых организмов.
4. создал учение о преобразовании биосферы в ноосферу, как результат действий человека.
111.Структура биосферы. Функции биосферы в развитии природы Земли и поддержания в ней динамических равновесий (окислительно-восстановительная, газообмен, концентрирование рассеянных в геосфере элементов, синтез и разложение органического вещества). Круговорот веществ.
Биосфера представляет собой многоуровневую систему, включающую подсистемы различной степени сложности. Границы биосферы определяются областью распространения организмов в атмосфере, гидросфере и литосфере (рис. 24.1). Верхняя граница биосферы проходит примерно на высоте 20 км. Таким образом, живые организмы расселены в тропосфере и в нижних слоях стратосферы. Лимитирующим фактором расселения в этой среде является нарастающая с высотой интенсивность ультрафиолетовой радиации. Практически все живое, проникающее выше озонового слоя атмосферы, погибает. В гидросферу биосфера проникает на всю глубину Мирового океана, что подтверждает обнаружение живых организмов и органических отложений до глубины 10—11 км. В литосфере область распространения жизни во многом определяет уровень проникновения воды в жидком состоянии — живые организмы обнаружены до глубины примерно 7,5 км.
Атмосфера. Эта оболочка состоит в основном из азота и кислорода. В меньших концентрациях она содержит углекислый газ и озон. Состояние атмосферы оказывает большое влияние на физические, химические и особенно биологические процессы на земной поверхности и в водной среде. Наибольшее значение для биологических процессов имеют кислород атмосферы, используемый для дыхания организмов и минерализации омертвевшего органического вещества, углекислый газ, расходуемый при фотосинтезе, а также озон, экранирующий земную поверхность от жесткого ультрафиолетового излучения. Вне атмосферы существование живых организмов невозможно. Это видно на примере лишенной жизни Луны, у которой нет атмосферы. Исторически развитие атмосферы связано с геохимическими процессами, а также жизнедеятельностью организмов. Так, азот, углекислый газ, пары воды образовались в процессе эволюции планеты благодаря (в значительной мере) вулканической активности, а кислород — в результате фотосинтеза.
Гидросфера. Вода является важной составной частью всех компонентов биосферы и одним из необходимых факторов существования живых организмов. Основная ее часть (95%) заключена в Мировом океане, который занимает примерно 70% поверхности Земного шара. Общая масса океанических вод составляет свыше 1300 млн. км3. Около 24 млн. км3 воды содержится в ледниках, причем 90% этого объема приходится на ледяной покров Антарктиды. Столько же воды содержится под землей. Поверхностные воды озер составляют приблизительно 0,18 млн. км3 (из них половина соленые), а рек—0,002 млн. км3.
Количество воды в телах живых организмов достигает примерно 0,001 млн. км3. Из газов, растворенных в воде, наибольшее значение имеют кислород и углекислый газ. Количество кислорода в океанических водах изменяется в широких пределах в зависимости от температуры и присутствия живых организмов. Концентрация углекислого газа также варьирует, а общее количество его в океане в 60 раз превышает его содержание в атмосфере. Гидросфера формировалась в связи с развитием литосферы, выделившей за геологическую историю Земли значительный объем водяного пара и так называемых ювенильных (подземных магматических) вод.
Литосфера. Основная масса организмов, обитающих в пределах литосферы, сосредоточена в почвенном слое, глубина которого обычно не превышает нескольких метров. Почвы, будучи, по терминологии В.И. Вернадского, биокосным веществом, представлены минеральными веществами, образующимися при разрушении горных пород, и органическими веществами — продуктами жизнедеятельности организмов.
Биотический круговорот. Главная функция биосферы заключается в обеспечении круговоротов химических элементов. Глобальный биотический круговорот осуществляется при участии всех населяющих планету организмов. Он заключается в циркуляции веществ между почвой, атмосферой, гидросферой и живыми организмами. Благодаря биотическому круговороту возможно длительное существование и развитие жизни при ограниченном запасе доступных химических элементов. Используя неорганические вещества, зеленые растения за счет энергии Солнца создают органическое вещество, которое другими живыми существами (гетеротрофами — потребителями и деструкторами) разрушается, с тем чтобы продукты этого разрушения могли быть использованы растениями для новых органических синтезов.
Важная роль в глобальном круговороте веществ принадлежит циркуляции воды между океаном, атмосферой и верхними слоями литосферы. Вода испаряется и воздушными течениями переносится на многие километры. Выпадая на поверхность суши в виде осадков, она способствует разрушению горных пород, делая их доступными для растений и микроорганизмов, размывает верхний почвенный слой и уходит вместе с растворенными в ней химическими соединениями и взвешенными органическими частицами в океаны и моря. Подсчитано, что с поверхности Земли за 1 мин испаряется около 1 млрд. т Н20 (на образование 1 г водяного пара необходимо 2,248 кДж). Энергия, затрачиваемая на испарение воды, возвращается в атмосферу (рис. 24.2). Циркуляция воды между Мировым океаном и сушей представляет собой важнейшее звено в поддержании жизни на Земле и основное условие взаимодействия растений и животных с неживой природой.
Под влиянием этого процесса происходит постепенное разрушение литосферы, перенос ее компонентов в глубины морей и океанов.
На создание органического вещества расходуется всего 0,1—0,2% солнечной энергии, достигающей поверхности планеты. Благодаря этой энергии осуществляется значительный объем работы по перемещению химических элементов.
В качестве примеров биотического круговорота рассмотрим круговороты углерода и азота в биосфере (рис. 24.3; 24.4). Круговорот углерода начинается с фиксации атмосферного диоксида углерода в процессе фотосинтеза. Часть образовавшихся при фотосинтезе углеводов используют сами растения для получения энергии, часть потребляется животными. Углекислый газ выделяется в процессе дыхания растений и животных. Мертвые растения и животные разлагаются, углерод их тканей окисляется и возвращается в атмосферу. Аналогичный процесс происходит и в океане.
Круговорот азота также охватывает все области биосферы (рис. 24.4). Хотя его запасы в атмосфере практически неисчерпаемы, высшие растения могут использовать азот только после соединения его с водородом или кислородом. Исключительно важную роль в этом процессе играют азотфиксирующие бактерии. При распаде белков этих микроорганизмов азот снова возвращается в атмосферу.
Показателем масштаба биотического круговорота служат темпы оборота углекислого газа, кислорода и воды. Весь кислород атмосферы проходит через организмы примерно за 2 тыс. лет, углекислый газ — за 300 лет, а вода полностью разлагается и восстанавливается в биотическом круговороте за 2 млн. лет (рис. 24.5).
Благодаря биотическому круговороту биосфере присущи определенные геохимические функции: газовая — биогенная миграция газов в результате фотосинтеза и азотфиксации; концентрационная — аккумуляция в своих телах живыми организмами химических элементов, рассеянных во внешней среде; окислительно-восстановительная — превращение веществ, содержащих атомы с переменной валентностью (например, Fе, Mn); биохимическая — процессы протекающие в живых организмах.
Стабильность биосферы. Биосфера представляет собой сложную экологическую систему, работающую в стационарном режиме. Стабильность биосферы обусловлена тем, что результаты активности трех групп организмов, выполняющих разные функции в биотическом круговороте,— продуцентов (автотрофы), потребителей (гетеротрофы) и деструкторов (минерализующие органические остатки) — взаимоуравновешиваются. То, что в биосфере поддерживается постоянство ее главных характеристик (гомеостаз), не исключает способности ее к эволюции.
Биофера включает живое вещество как совокупность жэивых организмов, биогенное вещество, которое создается, перерабатывается в процессе жизнедеятельности живых организмов : газы, нефть, каменный уголь, известники, сланцы. Косное вещество – образуется без участия живых организмов и абиогенных процессов. В биосферу входят радиоактивные вещества, рассеянные атомы, вещество космического происхождения.
Биосфера имеет несколько уровней. Границы бмиосферы – границы жизни на Земле. Ограничиваются температурой.
Верхняя граница – 20 км выше уровня земли. Тропосфера и нижний слой стратосферы. Нижняя – -20 км (глубина океана 10 км).
Между ними раполагаются литосфера, атмосфера и гидросфера.
Атмосфера включает в себя косное и биогенное вещество. Это важное условие жизни на Земле,т.к. регулирует климат, пропускает УФ-лучи, сохраняет тепло, служит средой для распространения света и звука, является источником кислорода для дыхания. Это среда, ерез которую Земля сообщается с Космосом.
Гидросфера – вся совокупность водных объектов ( почвенные воды. Ледники). Необходима, т.к. все организмы состоят из веществ, находящихся в водной фазе. Все биохимичекие процессы протекают в водной фазе.
Литосфера. Верхняя оболочка Земли. Самый верхний слой – почва. Толщина литосферы под горами составляет – 70-75, а под морем – 6-8 км. Почва – результата взаимодействия 5 факторов (по Докучаеву):
– геологические породы;
– климат;
– живые организмы;
– рельеф;
– время.
Через почву проходят обменные процессы энергией и веществами между живыми организмами, литосферой, гидросферой и атмосфкерой.
Живое вещество (о Вернадскому) – совокупность живых организмов. Биосфера – оболочка Земли, состав, энергетика, структура которой определяются деятельностью живых организмов. Вернадский доказал, что живые организмы, когда-то обитавшие и сейчас обитающие на Земле играют роль в геологической эволюции.
Живое вещество распространено неравномерно. В основно – в почве, на ее поверхности и верхней части гидросферы.
На суше выделяют 2 уровня жизни:
– на поверхности;
– в глубине.
В воде выделяют 3 уровня:
– планктон;
– нектон (плавающие организмы);
– бентос (на дне– глубинные существа).
Биомасса: на суше растений – 99,2%. Животных и микроорганизмов – 0,8%. В океане растений- 6,3%, животных и микроорганизмов – 93,7%,
По содержанию атомов водорода и кислорода живое вещество ближе к гидросфере, по содержанию углерода, кальция, азота – в живом веществе их концентрация выше, чем в гидросфере.
99% массы живых организмов пиходится на элементы земной коры.
112.Живое вещество биосферы. Количественная и качественная характеристика. Роль в природе планеты.
Живые организмы (живое вещество). В настоящее время описано около 300 тыс. видов растений и более 1,5 млн. видов животных. Из этого количества 93% представлено сухопутными, а 7% — водными видами животных. Суммарная биомасса организмов сухопутных видов образована на 99,2% зелеными растениями (2,4 • 1012 т) и на 0,8% животными и микроорганизмами (0,2 • 1011 т). В океане, напротив, на долю растений приходится 6,3% (0,2 • 109 т), а на долю животных и микроорганизмов — 93,7% (0,3 • 1010 т) совокупной биомассы. Несмотря на то что океан покрывает немногим более 70% поверхности планеты, в нем содержится лишь 0,13% биомассы всех живых существ, обитающих на Земле.
Расчеты показывают, что растения составляют около 21% всех учтенных видов. Однако на их долю приходится более 99% биомассы, тогда как вклад животных в биомассу планеты (79% видов) составляет менее 1%. Среди животных 96% видов приходится на долю беспозвоночных и только 4% на долю позвоночных, среди которых млекопитающие составляют примерно 10%.
Приведенные соотношения иллюстрируют фундаментальную закономерность организации биосферы: в количественном отношении преобладают формы, достигшие в процессе эволюции относительно низких степеней морфофизиологического прогресса.
Живое вещество по массе составляет 0,01—0,02% от косного вещества биосферы, однако играет ведущую роль в биогеохимических процессах благодаря совершающемуся в живых организмах обмену веществ. Так как субстраты и энергию, используемые в обмене веществ, организмы черпают из окружающей среды, они преобразуют ее уже тем, что в процессе своего существования используют ее компоненты.
Ежегодная продукция живого вещества в биосфере составляет 232,5 млрд. т сухого органического вещества. За это же время в масштабе планеты в процессе фотосинтеза синтезируется 46 млрд. тонн органических углеродсодержащих веществ. Для этого требуется, чтобы 170 • 109 т С02 прореагировало с 68 • 109 т Н20.
Таким образом, в результате фотосинтеза ежегодно образуется 115х х 109 т сухого органического вещества и 123 • 109 т 02. В течение года в процесс фотосинтеза вовлекаются также 6 • 109 т азота, 2 • 109 т фосфора и другие элементы, например калий, кальций, сера, железо. Приведенные цифры показывают, что живое вещество является наиболее активным компонентом биосферы. Оно производит гигантскую геохимическую работу, способствуя преобразованию других оболочек Земли в геологическом масштабе времени.
Живое вещество (о Вернадскому) – совокупность живых организмов. Биосфера – оболочка Земли, состав, энергетика, структура которой определяются деятельностью живых организмов. Вернадский доказал, что живые организмы, когда-то обитавшие и сейчас обитающие на Земле играют роль в геологической эволюции.
Живое вещество распространено неравномерно. В основно – в почве, на ее поверхности и верхней части гидросферы.
На суше выделяют 2 уровня жизни:
– на поверхности;
– в глубине.
В воде выделяют 3 уровня:
– планктон;
– нектон (плавающие организмы);
– бентос (на дне– глубинные существа).
Биомасса: на суше растений – 99,2%. Животных и микроорганизмов – 0,8%. В океане растений- 6,3%, животных и микроорганизмов – 93,7%,
По содержанию атомов водорода и кислорода живое вещество ближе к гидросфере, по содержанию углерода, кальция, азота – в живом веществе их концентрация выше, чем в гидросфере.
99% массы живых организмов пиходится на элементы земной коры.
Функции живого вещества.
1. Энергетическая.
2. Концентрационная.
3. Деструктивная.
4. Средообразующая.
5. Транспортная.
Энергетическая функция – поглощение солнечной энергии при фотосинтезе и химической энергии при хемосинтезе и передача энергии по пищевым цепям.
Концентрационная функция – способность накапливать химические элементы и использовать их при построении тела.
Деструктивная функция – ми нерализация неживого органического вещества и вовлечение образовавшихся веществ в биотический круговорот.
Средообразующая функция – преодалевание физико-химических параметров среды в результате синтеза и рнаспада.
Транспортная – перенос веществ против силы тяжести и в горизонтальном направлении.
Вернадский : «Живое вещество – самая активная форма материи во Вселенной».
Определение Вернадского.
Биосфера – комплексная геологическая оболочка, состоящая из атмо – , гидро – и литосферы, заселенная и преобразуемая живым веществом.»
Основой равновесия и устойчивости биосферы является круговорот веществ и энергии, а универсальной структурной единицей – биогеоценоз.
Сукачев разработал учение о биогеоценозе, 1947 год.
Биогеоценоз – растения, животные, микроорганизмы, находящиеся в постоянном контакте и взаимодействии с атмосферой, гидросферой, литосферой. Состоит из биотической части (биоценоз) и абиотической (биотоп).
Биоценоз – совокупноть популяций живых существ, населяю.щих определенную территорию.
Закон ограничивающего (лимитирующего) фактора или закон минимума Либиха — один из фундаментальных законов в экологии, гласящий, что наиболее значим для организма тот фактор, который более всего отклоняется от оптимального его значения. Поэтому во время прогнозирования экологических условий или выполнение экспертиз очень важно определить слабое звено в жизни организмов.
Именно от этого, минимально (или максимально) представленного в данный конкретный момент экологического фактора зависит выживание организма. В другие отрезки времени ограничивающим могут быть другие факторы. В течение жизни особи видов встречаются с самыми разными ограничениями своей жизнедеятельности. Так, фактором, ограничивающим распространение оленей, является глубина снежного покрова;[источник?]бабочки озимой совки (вредителя овощных и зерновых культур) — зимняя температура[источник?]и т. д.
Этот закон учитывается в практике сельского хозяйства. Немецкий химик Юстус Либих установил, что продуктивность культурных растений, в первую очередь, зависит от того питательного вещества (минерального элемента), который представлен в почве наиболее слабо. Например, если фосфора в почве лишь 20 % от необходимой нормы, а кальция — 50 % от нормы, то ограничивающим фактором будет недостаток фосфора; необходимо в первую очередь внести в почву именно фосфорсодержащие удобрения.
По имени учёного названо образное представление этого закона — так называемая «бочка Либиха». Суть модели состоит в том, что вода при наполнении бочки начинает переливаться через наименьшую доску в бочке и длина остальных досок уже не имеет значения.
113.Эволюция биосферы. Ресурсы биосферы.
Эволюция биосферы на Протяжении большей части ее истории осуществлялась под влиянием двух главных факторов: естественных геологических и климатических изменений на планете и изменений видового состава и количества живых существ в процессе биологической эволюции. На современном этапе в третичном периоде к ним присоединился третий фактор — развивающееся человеческое общество.
Этапы возникновения жизни, пути и механизмы ее эволюционного развития рассмотрены выше (см. гл. 1). Жизнь зародилась на Земле свыше 3,5 млрд. лет назад. Первыми живыми существами были анаэробы, которые получали энергию путем брожения. Так как брожение представляет собой относительно малопродуктивный способ энергообеспечения, примитивная жизнь не могла эволюционировать далее одноклеточной формы организации. Питание таких примитивных организмов зависело от опускавшихся на дно водоемов органических веществ, синтезируемых в поверхностных слоях воды абиогенным способом.
Недостаток органических веществ создал давление отбора, приведшее к возникновению фотосинтеза. Прогрессивное увеличение кислорода в воде за счет жизнедеятельности фотосинтезирующих организмов и его диффузии в атмосферу вызвало изменения в химическом составе оболочек Земли, прежде всего атмосферы, что в свою очередь сделало возможным и развитие более сложно организованных живых форм и быстрое распространение Жизни по планете. По мере увеличения содержания кислорода в атмосфере формируется достаточно мощный слой озона, защищающий поверхность Земли от проникновения жесткого ультрафиолетового излучения. В таких условиях жизнь смогла продвинуться к поверхности моря. Развитие механизма аэробного дыхания сделало возможным появление многоклеточных организмов. Примечательно, что первые такие организмы появились после того, как концентрация кислорода в атмосфере планеты достигла примерно 3%, что произошло около 600 млн. лет назад (начало кембрия).
Благодаря способности фотосинтезирующих морских организмов продуцировать такое количество кислорода, которое превышало потребности в нем обитателей планеты, стало возможным возникновение в процессе эволюции организмов более высокого уровня структурно-физиологической организации, их широкое расселение и проникновение Жизни в различные сферы обитания. В течение палеозойской эры живые существа не только заселили все моря, но и вышли на сушу. Развитие зеленых растений обеспечило образование больших количеств кислорода и органических веществ, что создавало благоприятные условия для последующей прогрессивной эволюции.
В середине палеозоя темпы потребления кислорода живыми организмами и расход его в абиотических процессах, а также темпы его образования сравнялись. Содержание кислорода в атмосфере начиная с этого периода истории Земли стабилизировалось на уровне примерно 20%.
С появлением человеческого общества в развитии биосферы намечается переход от биогенеза, обусловленного факторами биологической эволюции, к ноогенезу — развитию под влиянием разумной созидательной деятельности человечества.
Эволюция биосферы.
Этапы:
1. Аиогенный;
2. Период биогенеза;
3. Период ноогенеза.
Абиогенный период – 4 млрд. лет назад. Включает возникновение органических веществ из неорганических. Образование сахаров, аминокислот, азотистых оснований. Простейшего круговрота кислорода нет.
Период биогенеза – 3 млрд. лет назад. Характеризуется появлением анаэробных гетеротрофов, получающих энергию за счет брожения. Появляется жизнь и смерть, круговорота кислорода нет. Позднее (2 млрд. лет назад) – появляются фотосинтезирующие бактерий, 1% кислорода, первичная атмосфера. 1млрд.лет назад – появляются первые эукариоты, 3-4% кислорода. 600 млн. лет назад – 7% кислорода, появляются многоклеточные организмы. 400 млн. лет назад – накапливается кислород, происходит выход растений на сушу, протекает биологическая эволюция. Немного позденне появляется человек.
Период ноогенеза – решающий фактор эволюции – деятельностьчеловека.
– изменяет естественный рельеф, геологические и климатические условия;
– изменяет видовой состав живых существ.
Человек – мощнейшая геологическая сила на Земле.
Ноосфера – сфера разума и новый этап в развитии биосферы (Вернадский). Наука о ноосфере – ноогенетика.
Биосфера без человека существовала, а человек без нее – нет.
Ресурсы природы – явления, объекты, условия и процессы, используемые обществом для удовлетворения материальных, научных и культурных потребностей.
Природные ресурсы бывают:
1. Возобновимые ресурсы – ресурсы, находящиеся в пределах биосферного круговорота, способные к самовосстановлению за сроки, соизмеримые с хозяйственной деятельностью человека. К ним относятся: вода, кислород, почвы, растительность.
2. Невозобновимые ресурсы – не восстанавливаются. К ним относятся: минералы, нефть, природный газ, щебень, песок, гравий, видовой состав растений и животных.
3. Истощенные ресурсы – виды ресурсов, количество которых сократилось и дальнейшее их использование экономически нерационально. К ним относятся – озоновый экран, редкие виды растений и животных, воды Арала.
114.Международные и национальные программы по изучению биосферы.
В настоящее время все ресурсы задействованы. Нерациональное природопользование ведет к экологическому кризису. Преодоление экологического кризиса осуществляется по закону РФ «Об охране окружающей среды» 1991 года. Этот закон предлагает основные направления выхода из экологического кризиса.
1. Приоритет охраны жизни и здоровья человека.
2. Сочетание экологических и экономических интересов.
3. Рациональное использование природных ресурсов.
4. Платность природопользования.
5. Международное сотрудничество.
Направления выхода из экологического кризиса РФ:
1) совершенствование технологий, создание экологически технологий, внедрение безотходных производств;
2) развитие экономического механизма охраны среды (штрафы);
3) применение юридической ответственности за экологические правонарушения;
4) формирование экологического мышления;
5) совершенствование международных отношений, международно-правового направления.
Международное сотрудничество.
1. Стокгольмская конференция ООН 1972 года провозгласила направления действий мирового сообщества в области охраны окружающей среды и объявила 5 июня всемирным днем охраны окружающей среды.
2. Всемирная Хартия Природы 1982 года. Принята концепция устойчивого развития общества.
§
Закон РФ 1995год.
Территории предназначены для поддержания экологического баланса, сохранения генетического разнообразия, эволюции экосистем, изучения влияния на них антропогенных факторов.
К ним относят:
– заповедники, в т.ч. биосферные;
– национальные парки;
– заказники;
– памятники природы;
– ботанические сады.
Заповедники – участки территорий, которые полностью изъяты из хозяйственного использования.
На 1997 год в РФ было 95 заповедников.
Биосферные заповедники используются в качестве заповедно-эталонных объектов. В мире существует единая сеть из 300 биосферных заповедников, в РФ их – 11. они работают по программе ЮНЕСКО.
Национальные парки – большие природные территории и акватории, где обеспечивается поддержание экологического баланса, сохранение природных систем, регуляция туризма и отдыха, разработка научных методов сохранения природного комплекса в условиях массового. В РФ – 33.
Заказники – территории, созданные на определенный срок для восстановления природных комплексов. В России 1600. в заказниках восстанавливают плотность популяций.
Памятники природы – уникальные невоспроизводимые природные объекты, к ним относятся:
– пещеры;
– вековые деревья;
– водопады.
В России 8000.
Ботанические сады – коллекции деревьев и кустарников, созданные искусственным путем для сохранения биологического разнообразия.
117.Определение науки экологии. Возникновение и основные этапы развития экологии. Междисциплинарный характер экологии.
118.Среда как экологическое понятие. Факторы окружающей среды, их классификация, взаимодействие и воздействие на экологические системы.
119.Экосистема, биогеоценоз, антропобиоценоз. Сукцессия экосистемы и её этапы.
120.Экология человека. Структура и содержание экологии человека. Основные её подразделения: эндоэкология, аутэкология, демэкология и синэкология человека.
121.Ноосфера. Представления о ноосфере. Медико-биологические аспекты ноосферы.
122.Морфофизиологическая характеристика людей естественных экосистем и географических районов – зоны тропиков, высокогорья, аридных областей, Арктики и континентальной Сибири, зоны умеренного климата).
123.Медицинская экология. Появление нового типа заболеваний человека- экологически зависимых болезней. Биогеохимические провинции и экологические заболевания человека
124.Основные формы биотических связей в природе. Паразитизм как биологический феномен, его особенности как формы межвидовых взаимодействий. Классификация паразитизма и паразитов. Пути происхождения экто – и эндопаразитов. Геогельминты и биогельминты.
Антибиоз — невозможность сосуществования двух видов организмов, основанная на конкуренции прежде всего за источники питания. Примером служат взаимоотношения сапрофитных бактерий и ряда плесневых грибов. Первые способны быстро заселять среды, богатые органическими веществами, за счет интенсивного размножения, а вторые, значительно уступая им в этом, приобрели способность делать субстрат неблагоприятным для жизнедеятельности бактерий, выделяя в него продукты своего метаболизма — антибиотики. В результате среда используется либо грибами, либо бактериями, успевшими попасть в нее и размножиться раньше.
Симбиоз в переводе с греческого означает «сожительство». Формы симбиоза разнообразны. В некоторых случаях отношения между организмами разных видов являются взаимополезными настолько, что раздельное их существование вообще невозможно. Такой симбиоз называют мутуализмом. Примером мутуалистических взаимоотношений является сожительство человека с микрофлорой его кишечника, основным компонентом которой являются разнообразные штаммы бактерий кишечной палочки Escherichia coli.
Бактерии в таком сожительстве находят благоприятную среду обитания и неисчерпаемый источник питания. Нормальное же пищеварение в кишечнике человека и всасывание рядов витаминов возможно только при участии бактерий. После длительного лечения больных различными инфекционными заболеваниями с помощью антибиотиков у них нередко наряду с подавлением жизнедеятельности болезнетворных бактерий наблюдается состояние дисбактериоза — гибель нормальных бактерий кишечника и усиленное размножение бактерий, нечувствительных к антибиотику, и микроскопических грибов, которые в свою очередь сами могут явиться причиной заболевания. Для восстановления нормальной кишечной микрофлоры часто необходимым является искусственное заселение пищеварительной системы человека симбионтными штаммами кишечной палочки.
Комменсализм — форма симбиоза, при которой один вид использует остатки или излишки пищи другого, не причиняя ему видимого вреда. Часто комменсалы даже поселяются в теле хозяина, не снижая его жизнеспособности. Примером комменсалов являются непатогенные ротовая и кишечная амебы, живущие в пищеварительной системе человека и питающиеся бактериями.
При хищничестве между организмами разных видов существуют только пищевые взаимоотношения, а пространственные отсутствуют. Хищники используют представителей другого вида для питания однократно, убивая их.
Наибольшее значение для медицины имеет паразитизм — форма межвидовых взаимоотношений, при которой один вид использует другой как источник питания и среду обитания. В тех случаях, когда паразит не живет в организме хозяина, он посещает его для питания многократно.
Слово паразит стало применяться в Древней Греции для обозначения пассивных участников жертвоприношений во время религиозных обрядов. Позже паразитами стали называть непрошеных гостей, а также персонажей драматических произведений, не выполняющих в действии серьезных функций (рис. 18.2). Понятие «паразит» в современном биологическом смысле стало применяться в Европе с XV-XVI вв.
Разные формы симбиотических взаимоотношений организмов не являются абсолютно стойкими и могут переходить друг в друга. Так, комменсалы могут становиться паразитами при ослаблении иммунитета хозяина. При нормальном питании и физическом здоровье хозяина некоторые паразиты могут долгое время не оказывать на него патогенного действия. Некоторые хищники, питающиеся мелкими животными, могут становиться паразитами крупных, и наоборот. Об этом подробнее см. разд. 18.5.
Условность классификации биотических связей и нечеткость их отграниченности друг от друга являются отражением эволюции не только взаимодействующих видов, но и самих межвидовых взаимоотношений.
КЛАССИФИКАЦИЯ ПАРАЗИТИЗМА
И ПАРАЗИТОВ
Формы паразитизма чрезвычайно многообразны, и классификация их возможна по разным основаниям. С точки зрения обязательности паразитического образа жизни для данного вида различают истинный и ложный, а также облигатный и факультативный паразитизм.
При истинном паразитизме взаимоотношения между паразитом и хозяином являются закономерными и имеют эволюционную основу. Паразитология изучает в основном феномен истинного паразитизма.
Ложный паразитизм — явление для данного вида случайное. В нормальных условиях данный вид ведет свободный образ жизни. При попадании в организм хозяина ложный паразит может некоторое время сохранять жизнеспособность и нарушать жизнедеятельность хозяина. Примерами ложного паразитизма являются случаи обнаружения пиявок в носовой полости и носоглотке человека. Ложный паразитизм пиявок может привести хозяина к смерти в связи с закупоркой дыхательных путей или из-за носовых кровотечений, которые они могут вызвать.
Облигатный паразитизм — паразитизм, являющийся обязательным для данного вида организмов. Абсолютное большинство видов паразитов относятся к этой группе.
Факультативные паразиты способны вести свободный образ жизни, но, попадая в организм хозяина, проходят в нем часть цикла своего развития и нарушают его жизнедеятельность. Таковы многие виды синантропных мух, личинки которых могут нормально развиваться либо в пищевых продуктах человека, либо в его кишечнике, вызывая кишечный миаз (см. разд. 21.2.4).
По времени контакта хозяина и паразита паразитизм бывает временным и постоянным. Временные паразиты обычно посещают хозяина только для питания. Это в основном кровососущие членистоногие. Постоянные паразиты подразделяются на стационарных и периодических.
Стационарные паразиты всю жизнь проводят на хозяине или внутри него. Примерами являются вши, чесоточный клещ, трихинелла спиральная и многие другие. Периодические паразиты часть своего жизненного цикла проводят в паразитическом состоянии, остальное время обитают свободно. Типичным паразитом такого рода является угрица кишечная.
Нередко паразитический образ жизни ведут только личинки, в то время как половозрелые формы являются свободноживущими. Паразитизм такого рода называют ларвальным (личиночным). Примерами служат вольфартова муха, оводы и др. (см. разд. 21.2.4). Противоположное явление, когда паразитом является половозрелая форма, а личинка обитает в открытой природе, называют имагинальным паразитизмом. К паразитам этого типа относят, например, анкилостомид, личинки которых живут в почве, а взрослые стадии — в двенадцатиперстной кишке человека.
Особенно большое медицинское значение имеет классификация паразитов по их локализации в организме хозяина. Эктопаразиты находятся на покровах хозяина. К ним относят кровососущих насекомых и клещей. Эндопаразиты обитают внутри хозяина. Их подразделяют на паразитов, обитающих в полостных органах, связанных с внешней средой (пищеварительная, дыхательная и мочеполовая системы), и паразитов тканей внутренней среды (опорно-двигательный аппарат, система крови, соединительная ткань). Примерами первых являются аскарида, легочный сосальщик, урогенитальная трихомонада, вторых — ришта, малярийный плазмодий, лейшмании.
Любой подход к классификации паразитизма не дает возможность строго разграничить формы этого сложного экологического явления. Многие виды на протяжении жизненного цикла могут быть по отношению к разным хозяевам и ларвальными, и имагинальными паразитами. Так, сосальщики на начальных этапах развития ведут свободный образ жизни. Позже их личинки обитают в промежуточном хозяине, затем вновь образуются свободноживущие личинки, которые, обнаружив второго промежуточного или окончательного хозяина, паразитируют у него на половозрелой стадии.
В процессе жизнедеятельности нередко паразиты осуществляют миграцию в организме хозяина и способны таким образом вначале обитать в полостных органах, а затем перемещаться в ткани внутренней среды. Таковы трихинелла и свиной цепень. Возможен переход от эктопаразитизма к паразитированию в тканях внутренней среды. К таким видам относятся, например, личинки вольфартовой мухи.
Отсутствие четких границ между разными формами паразитизма отражает объективную ситуацию — эволюцию этого экологического феномена.
Своеобразной экологической группой паразитов являются сверхпаразиты. В качестве среды обитания и источника питания ими используются другие паразитические организмы. Обычно сверхпаразиты еще более мелкие и низко организованы, чем паразиты (рис. 18.3). Они могут поражать как простейших, так и многоклеточных паразитов. Сверхпаразитизм — очень широко распространенное явление. Так, подсчитано, что только один вид свободноживущих бабочек — луговой мотылек Loxostege sticticalis — является хозяином 40 видов паразитов, за счет которых существуют еще 12 видов сверхпаразитов. Среди сверхпаразитов, обитающих в паразитах человека, известны несколько видов микроспоридий, относящихся к классу споровиков и встречающихся в цитоплазме балантидия (см. разд. 19.2.3), в клетках паренхимы цепней (см. разд. 20.1.2) и в гонадах аскарид (см. разд. 20.2.1).
Сверхпаразиты имеют огромное экологическое значение, выполняя функции стабилизаторов численности популяций паразитов. Медицинское значение сверхпаразитов еще не изучено, но не исключено, что и в популяциях человека они могут выполнять роль факторов, сдерживающих численность паразитов.
Явление паразитизма, как и любой другой экологический феномен, возникло разными путями. С одной стороны, по-разному развиваются взаимные адаптации паразитов и хозяев в разных систематических группах организмов — классах и типах, с другой — различны направления эволюции, ведущие к возникновению разнообразных форм паразитизма.
Первый подход к исследованию происхождения паразитизма конкретен. Он рассматривается при изложении материала по частной паразитологии в разделах, посвященных описанию характеристик типов и классов паразитических организмов и их экологических групп. Второй подход вскрывает общие закономерности перехода к паразитическому существованию вне зависимости от систематического положения организмов, занимающих новые экологические ниши.
Наиболее просто объясняется происхождение эктопаразитизма. Один из путей к этому — через увеличение количества источников питания с последующей их сменой. Так, многие насекомые имеют колюще-сосущий ротовой аппарат, питаясь соками растений. Но питание за счет прокалывания ткани и всасывания жидкости и есть способ поглощения пищи всеми кровососущими членистоногими, ряд которых, потребляя кровь человека и теплокровных животных, продолжает пользоваться также и соками растений, s
Другой путь, ведущий к эктопаразитизму, — хищничество. Активные хищники, осваивающие для питания все более крупные жертвы, становятся вначале временными, а затем и постоянными эктопаразитами за счет удлинения контактов с организмом хозяина. Так, многие пиявки, ведущие себя как хищники по отношению к мелким организмам, становятся паразитами более крупных животных, питаясь их кровью. Увеличение продолжительности питания — основное направление перехода от временного к постоянному эктопаразитизму. Действительно, из большого количества кровососущих форм членистоногих наиболее длительное питание на хозяине характерно именно для постоянных паразитов, степень контакта которых с хозяевами наиболее высока.
Иной путь возникновения эктопаразитизма — через усиление контакта так называемых гнездовых паразитов с поверхностью тела хозяина. Животные, обитающие в убежище другого вида, могут питаться его перьями, волосами и отпадающими чешуйками кожного эпидермиса. Переход к постоянному обитанию на поверхности тела хозяина дает паразиту большие преимущества. Возможно, так возник паразитизм пухоедов, власоедов птиц и млекопитающих и группы клещей — обитателей эпидермиса животных и человека.
Основная масса случаев эндопаразитизма в полостных органах, имеющих связь с внешней средой, представляет собой явление, развившееся в результате случайного заноса в организм цист, яиц или личинок свободноживущих видов, предварительно имеющих адаптации к обитанию в почве или в воде, содержащей избыток органического вещества. Примером является угрица кишечная, которая в своем развитии сохранила возможность обитать и размножаться как в почве, так и в организме человека (см. разд. 20.2.1.1).
Возможен переход к паразитированию в одном хозяине после предварительной адаптации к обитанию в другом, служащем источником питания первого. Так, известен целый ряд гельминтов, которые, обитая в кишечнике рыбы, не перевариваются в пищеварительной системе хищников, съевших паразитов вместе с хозяином и продолжающих паразитировать в кишечнике или тканях уже нового вида (см. разд. 20.1.2.1).
Не исключается и вариант перехода к полостному паразитизму видов, предварительно адаптированных к эктопаразитизму. Этим путем, вероятно, эволюционируют некоторые насекомые, большую часть цикла развития проводящие в ротовой полости птиц, но выходящие для размножения на перьевой покров их головы.
Наиболее сложно и многообразно происхождение паразитов тканей внутренней среды. Один из путей — через изменение инстинкта откладки яиц и предварительных адаптации к эктопаразитизму. Таким путем, вероятно, произошел тканевой паразитизм личинок мух и оводов (см. разд. 21.2.4), откладывающих яйца на поверхности кожи и слизистых оболочек животных и человека. Личинки при этом вскоре погружаются под покровы и ведут типичный эндопаразитический образ жизни.
Многие паразиты приспособились к обитанию в тканях после освоения полостных органов, связанных с внешней средой. Так, по-видимому, шла эволюция паразитизма у ленточных червей, у трихинеллы спиральной (см. разд. 20.2.1.2). В цикле развития этих паразитов имеются формы, обитающие как в кишечнике, так и в тканях.
Некоторые паразиты внутренней среды возникли, вероятно, предварительно адаптировавшись к обитанию в пищеварительной системе членистоногих, а с переходом последних к гематофагии заселили новую и труднодоступную экологическую нишу — кровь и другие ткани мезодермального происхождения.
Таким образом, путей перехода к паразитизму у разных видов животных много, но несомненным остается одно: паразитизм — явление вторичное. Об этом свидетельствует наличие в жизненных циклах многих, даже наиболее специализированных паразитов, свободноживущих стадий, рекапитулирующих свободный образ жизни предков (см., например, разд. 20.1.1).
Живые организмы находятся в биотическом окружении, поэтому они оказывают воздействие друг на друга. Между организмами разных биологических видов устанавливаются связи в результате эволюции. Одни организмы живут отдельно, а другие приобрели приспособления для совместной жизни. Одни использовали других – квартиранство.
 Паразитизм и мутуализм возникли в ходе эволюции отдельно друг от друга, поэтому не переходят друг в друга.
Паразитизм и мутуализм возникли в ходе эволюции отдельно друг от друга, поэтому не переходят друг в друга.
Догель, 1947 г. « Паразиты – это такие организмы, которые используют другие организмы в качестве источника пищи и среды обитания, возлагая при этом частично или полностью на своих хозяев задачу регуляции своих взаимоотношений с окружающей средой».
Среда обитания паразита – другой организм – хозяин. Организм хозяина имеет собственный гомеостаз, защитные механизмы от всего инородного. Паразит – чужеродный организм. В процессе эволюции у паразита выработались приспособления для существования в живом организме, но хозяин также приспособился к паразиту. В процессе эволюции происходит соревнование. Паразиты – процветающие виды, имеющие биологический прогресс. Они очень изменчивы, пластичны. Паразиты – очень древние виды. Паразитизм существует с момента зарождения жизни (например, бактериофаги). У паразитов много упрощений (нет многих ферментов, пищеварительной трубки, у плоских червей пристеночное переваривание, как в тонком кишечнике). Поселяясь, в организме хозяина, они оказывают на него патогенное действие, зависящее от:
– размеров;
– морфологии;
– физиологии;
– интенсивности инвазии;
– локализации в организме хозяина;
– особенностей инвазии во время заражения.
Инвазия – количество паразитов в теле одного хозяина.
Болезни, вызываемые организмами животной природы – инвазионные.
Болезни, вызываемые растительными и другими организмами – инфекционные.
После поселения в организме хозяина паразит может воздействовать на его организм.
Геогельминты:
У этих червей яйца или личинки обязательно развиваются в поверхностных слоях почвы при доступе кислорода и достаточной влажности. За исключением острицы, все гельминты встречаются чаще в регионах с жарким и влажным климатом, который предоставляет личинкам и яйцам больше возможностей развиваться в почве. В Арктике и южных приполярных областях геогельминты не встречаются. Самцы и самки гельминтов легко различимы: самцы большинства видов имеют загнутый на брюшную сторону или спирально закрученный задний конец тела, в то время как у самок он прямой.
Геогельминты обитают в просвете кишки и размножаются яйцами, которые выводятся с фекалиями и развиваются далее в почве. Они либо сами через определенное время становятся инвазионными, либо из них развиваются личинки, ведущие некоторое время свободный образ жизни и позже становящиеся инвазионными. Геогельминты, поражающие человека, не могут паразитировать у животных. Соответственно этому нематодозы, вызываемые этими паразитами, являются антропонозными болезнями. Заражение большей частью геогельминтов осуществляется при проглатывании яиц или личинок с продуктами, загрязненными почвой.
Для диагностики всех нематодозов этой группы важно обнаружение яиц в фекалиях больного.
Профилактические меры направлены на предотвращение попадания инвазионных яиц в пищеварительную систему—личная гигиена и гигиена питания, реже — другие меры. Часть геогельминтов, попадая в пищеварительную систему человека, быстро достигают половой зрелости и начинают размножаться в кишечнике, не мигрируя по организму хозяина. Личинки других перед достижением половой зрелости обязательно передвигаются по кровеносным сосудам и дыхательной системе и только после этого развиваются в кишечнике.
Биогельминты:
Все нематоды этой группы, поражающие человека, живородящи и большую часть цикла развития проводят у человека в тканях внутренней среды. Промежуточные хозяева их очень разнообразны — от циклопов и насекомых до медведей и человека. Окончательными хозяевами могут быть различные дикие и домашние животные, поэтому заболевания, которые вызывают эти паразиты, относятся к разряду природно-очаговых. Для попадания в места окончательной локализации биогельминты осуществляют миграцию по лимфатическим и кровеносным сосудам. Кроме того, они особенно активно взаимодействуют с иммунной системой хозяина. Поэтому в клинической картине нематодозов-биогельминтозов ведущими симптомами являются ток-сико-аллергические реакции. Важно также механическое и местное токсическое действие. Диагноз биогельминтозов часто затруднен. Приходится прибегать к методам биопсии и иммунологическим реакциям. Профилактика зависит от путей заражения, которые бывают различны.
125.Паразитоценоз. Взаимоотношения в системе паразит-хозяин на уровне отдельной особи. Адаптации к паразитическому образу жизни. Факторы действия паразита на организм хозяина.
Паразитоценоз (от паразиты и греческого koinós — общий), совокупность паразитов, обитающих в каком-либо органе, в непосредственно связанных друг с другом органах или во всём организме животного или человека. Понятие П. введено в науку советским учёным Е. Н. Павловским.В состав П. могут входить паразитические животные, бактерии, грибы, вирусы (независимо от того, вызывают они заболевание хозяина или нет). Видовой состав П. и количественные соотношения входящих в П. видов сильно варьируют у разных животных-хозяев, даже относящихся к одному виду. Наиболее полное определение видового состава П. необходимо для познания сложных взаимоотношений П. в целом, а также отдельных его компонентов с организмом хозяина, что позволяет правильно понимать явление паразитоносительства, сущность паразитарных и инфекционных болезней и избирать методы действенного их лечения и профилактики.
АДАПТАЦИИ К ПАРАЗИТИЧЕСКОМУ ОБРАЗУ ЖИЗНИ. ОСНОВНЫЕ ТЕНДЕНЦИИ
Переход к паразитическому образу жизни сопровождается появлением у паразитов ряда адаптации, облегчающих их существование, развитие и размножение в специфических условиях организма хозяина Разнообразие форм паразитизма, различное систематическое положение паразитов (их принадлежность к разным отрядам, классам и типам), а также обитание их в разных органах и системах хозяина обусловливают многообразие этих адаптации.
Однако некоторые приспособления являются абсолютно универсальными. К ним в первую очередь относятся высокая плодовитость и особенности половой системы. Действительно, возможность оставления потомства и попадания его в благоприятную среду — организм хозяина — у паразитов часто ничтожна. В связи с этим интенсивность размножения паразитов по сравнению со свободноживущими формами гораздо более велика. Достигается это разными способами. У многоклеточных это сильная степень развития половой системы и образование огромного количества половых продуктов. Этому способствуют первичный гермафродитизм плоских червей, изначально высокая плодовитость круглых червей и основной массы членистоногих. Нередко высокая интенсивность полового размножения дополняется размножением личиночных стадий жизненного цикла. Особенно это характерно для сосальщиков, личинки которых размножаются партеногенетически, а у некоторых ленточных червей — внутренним или наружным почкованием.
Некоторые паразиты из типа простейших приобретают способность к множественному делению — шизогонии, когда из одного паразита может образоваться более 1000 дочерних особей, или к спорогонии, в результате которой из одной особи могут образоваться десятки тысяч организмов следующего поколения. Практически у всех эктопаразитов и паразитов, обитающих в полостных органах, имеются адаптации для прикрепления к телу хозяина. Они встречаются у простейших (присасывательные диски лямблии), у гельминтов (присоски, шипики, крючья плоских червей, хитинизированный ротовой аппарат ряда круглых червей) и паразитических членистоногих (своеобразные конечности) (рис. 18.4).
Эндопаразиты, обитающие в полостных органах, имеют покровы, обладающие антиферментными свойствами, быстро регенерирующие либо вообще непроницаемые для ферментов хозяина. Паразиты, живущие в тканях, часто там инкапсулируются (рис. 18.5).
Паразиты, питающиеся кровью (представлены в основном членистоногими), имеют колюще-сосущий ротовой аппарат, а также сильно растяжимый хитиновый покров, часто разветвленную пищеварительную трубку (рис. 18.6), антикоагулянтные свойства слюны и консервантные свойства ферментов пищеварительной системы.
Эндопаразиты, активно отыскивающие хозяина, обладают органами ориентации в среде, используемыми для поисков хозяина (светочувствительные глазки, термо- и хеморецепторы), и органами передвижения (рис. 18.7).
Передний конец тела паразитов, внедряющихся в организм хозяина, снабжен органами проникновения — специализированными железами, колющими стилетами и т.д. Это касается даже некоторых простейших, способных проникать в ткани хозяина через неповрежденные покровы.
Все паразиты, развивающиеся со сменой хозяев, используют в качестве таковых виды, связанные между собой непосредственными пищевыми взаимоотношениями или обитающие с ними в одной среде. Большинство промежуточных хозяев являются источником питания для основных. Другой распространенный путь попадания паразита в организм хозяина — это использование многочисленных переносчиков, которые обеспечивают не только постоянную циркуляцию паразитов в экологических системах, но и их широкое расселение.
Высшей степенью адаптации паразитов к хозяевам является наблюдаемая часто полная зависимость паразита от жизнедеятельности хозяев. При этом паразит нередко вызывает такие реакции хозяина, которые обеспечивают максимальную вероятность заражения последнего. Так, самки остриц, откладывая яйца в области анального отверстия, вызывают зуд. Расчесывание зудящих мест способствует распространению яиц этого паразита руками по окружающим предметам. Таким же образом обеспечивается расселение чесоточного клеща. Зуд в пораженной конечности, прекращающийся от соприкосновения с водой, способствует циркуляции в природе такого паразита, как ришта. Высокая температура больных паразитарными заболеваниями, распространяющимися с помощью кровососущих членистоногих, привлекает переносчиков нередко с больших расстояний и также оказывается полезной для паразитов.
Нередко особенности жизнедеятельности паразитов оказываются синхронизированными с образом жизни хозяев. Так, откладка яиц шистосомами происходит обычно в самое жаркое время суток, когда наиболее вероятным оказывается контакт хозяев с водой, куда для развития должны попасть яйца этих паразитов. В это же время в поверхностных слоях воды скапливаются в поисках хозяев церкарии этих шистосом. Таким образом облегчается циркуляция паразита сразу на двух стадиях его жизненного цикла (см. разд. 20.1.1.2). Если в циркуляцию паразита включены несколько хозяев, то наблюдаются их взаимные адаптации, оказывающиеся выгодными паразитам и обеспечивающие его эффективное развитие. Так, выход микрофилярий в кровеносные сосуды человека происходит в часы суток, соответствующие периоду максимальной активности кровососущих насекомых, являющихся их переносчиками (см. разд. 20.2.1.2).
Нередко паразиты даже модифицируют поведение одних хозяев таким образом, что в результате облегчается их попадание к другим. Так, рыбы, пораженные личинками ленточных червей, плавают в основном у поверхности воды и чаще вылавливаются рыбаками и хищными животными. Ленточные черви, использующие в качестве промежуточных хозяев копытных животных, снижают их жизнеспособность, и, таким образом, хищники поедают их в первую очередь. Сосальщики, заражающие травоядных животных и человека через случайное проглатывание насекомых, вызывают обездвиживание последних и плотное прикрепление к растениям, облегчающее им попадание в пищеварительный тракт хозяина (см. раздел 20.1.1.3. стр. 245).
Одновременно с перечисленными признаками свойства паразитов переживать неблагоприятные условия внешней среды являются также несомненными адаптациями к паразитизму. Большинство простейших, заражение которыми происходит без участия переносчиков, во внешней среде способны инцистироваться. Яйца большинства гельминтов обладают феноменальной устойчивостью к неблагоприятным воздействиям. Капсулы с личинками трихинелл переносят не только промораживание, кипячение, но и многократное прохождение через пищеварительную систему рыб, земноводных, птиц, насекомых и ракообразных, не теряя жизнеспособности.
Для большинства паразитов, переживших нахождение во внешней среде или в промежуточном хозяине в покоящихся стадиях и попавших в организм окончательного хозяина, существует комплекс условий, являющийся сигналом к началу активной жизнедеятельности. У млекопитающих он часто неспецифичен: это температура тела около 37°С, водная среда и высокая ее кислотность, а также наличие ферментов желудочного сока. Такие условия характерны для желудка любого млекопитающего, поэтому цисты, яйца и другие инвазионные стадии паразитов, обитающие у разных млекопитающих могут, попадая в желудок человека, начинать развитие. Некоторые из них проходят лишь часть цикла и, не находя специфических условий, гибнут (см. разд. 20.2.1.3), успевая, однако, привести к тяжелым последствиям.
Указанные особенности паразитов, общие для многих из них, не состоящих в родстве, возникли в разных их группах независимо друг от друга, иллюстрируя конвергентный характер эволюции организмов разных видов, классов и типов, адаптирующихся к сходным условиям.
Факторы действия паразитов:
· Механический (механическое разрушение органов путем давления, внедрения, прикрепления).
· Химический (отравление организма хозяина ядовитыми продуктами распада паразита после его смерти).
· «Отнятие пищи» (непосредственное отнятие пищевых веществ и витаминов у хозяина). Например, лентец широкий отнимает витамин В12/
Когда паразит вступает в контакт с хозяином, постепенно формируется система паразит – хозяин. Наступает взаимная адаптация. Если не приспособятся, наступает гибель. В результате взаимодействия формируется энергетически сбалансированная система, пищи хозяина хватает для обоих. Иногда вес хозяина увеличивается. Паразит активно изменяет гомеостаз хозяина. Паразит стремится создать такие условия, которые будут для паразита оптимальными. У него есть вещества, изменяющие обмен веществ хозяина. Паразит выделяет биологически активные вещества, которые повышают обмен веществ хозяина, при этом продолжительность жизни хозяина снижается.
126.Циклы развития паразитов. Чередование поколений и феномен смены хозяев. Основные, резервуарные и промежуточные хозяева. Расселение паразитов и проблемы поиска хозяина.
Онтогенез паразитов обычно бывает сложнее, чем развитие свобод-ноживущих видов. Действительно, свободноживущие организмы довольно легко преодолевают проблемы размножения и расселения, что значительно усложнено у паразитов. Поэтому большинство паразитов нередко развиваются со сложным метаморфозом, включающим много личиночных стадий, обитающих в разных средах и выполняющих разные функции: расселения, активного роста, пассивного ожидания попадания в другую среду обитания и иногда даже размножения.
Совокупность всех стадий онтогенеза паразита и путей передачи его от одного хозяина к другому называют его жизненным циклом. Личинки могут вести как свободный, так и паразитический образ жизни. Хозяин, в котором обитают личинки паразита, носит название промежуточного. Значение промежуточных хозяев в циклах развития паразитов очень велико: они являются источниками заражения окончательных хозяев, часто выполняют расселительные функции, а иногда обеспечивают выживание популяций паразита в случае временного исчезновения окончательных хозяев.
Иногда в цикле развития паразита последовательно сменяются два-три промежуточных хозяина и даже больше. Хозяина, в котором развивается и размножается половым путем половозрелая стадия паразита, называют окончательным или дефинитивным. Заражение его осуществляется либо при поедании промежуточного хозяина, либо при контакте с последним в одной среде обитания.
Выделяют также понятие «резервуар паразита», или «резервуарный хозяин». Это такой хозяин, в организме которого возбудитель заболевания может жить долго, накапливаясь, размножаясь и расселяясь по окружающей территории.
Наиболее часто резервуарами паразитов служат их дефинитивные хозяева. В том случае, когда продолжительность жизни промежуточного хозяина велика, а личинка в нем долго сохраняет жизнеспособность и иногда даже размножается, он также может выполнять роль резервуара. Продолжительность жизненного цикла разных паразитов очень сильно колеблется в зависимости от их систематического положения, видовой принадлежности и условий. Так, жизнь аргасовых клещей может продолжаться до 20 лет, кровяных сосальщиков — до 40, а детская острица и карликовый цепень живут не более 2 мес. Знание длительности онтогенеза паразитов необходимо для разработки мер профилактики паразитарных заболеваний.
Расселение паразитов может происходить на разных стадиях их жизненного цикла. Расселение во времени обычно осуществляется покоящимися стадиями: развитие на этих стадиях приостанавливается до тех пор, пока не возникают новые условия, благоприятные для дальнейшего развития. Такими стадиями у простейших являются цисты, а у гельминтов — обычно яйца и иногда инкапсулированные личинки. Обычно покоящиеся стадии очень устойчивы к изменениям внешней среды. Так, яйца аскариды могут сохраняться жизнеспособными до 7 лет, а цисты дизентерийной амебы — до 7 мес. При попадании покоящейся стадии в благоприятного хозяина перемещение последнего способствует расселению паразита (часто далеко за пределы ареала его первоначального существования). Цисты, яйца и инкапсулированные личинки могут также разноситься ветром, водными потоками и животными — механическими переносчиками.
§
К типу Простейшие относят организмы, тело которых состоит из одной клетки, функционирующей, однако, как целый организм. Клетки простейших способны к самостоятельному питанию, передвижению, защите от врагов и к переживанию неблагоприятных условий. В строении простейших обнаруживаются как все особенности эукарио-тических клеток, так и специфические органеллы, обеспечивающие выполнение организменных функций.
Питание простейших происходит с помощью пищеварительных вакуолей, содержащих пищеварительные ферменты и связанных по происхождению с лизосомами. Оно осуществляется за счет фаго- или пиноцитоза. Остатки непереваренной пищи выбрасываются наружу. Некоторые простейшие содержат хлоропласты и способны питаться за счет фотосинтеза.
Большинство простейших имеют органеллы передвижения: жгутики, реснички и псевдоподии (временные подвижные выросты цитоплазмы). Формы органелл движения лежат в основе систематики простейших.
Пресноводные свободноживущие простейшие имеют органеллы, регулирующие водно-солевой баланс, — сократительные вакуоли. Периодически они сокращаются и выделяют во внешнюю среду избытки воды и жидкие продукты диссимиляции. Морские и паразитические простейшие, живущие в среде с высокой концентрацией солей, могут не иметь сократительных вакуолей.
Размножение простейших осуществляется обычно разными формами деления — разновидностями митоза. Характерен также половой процесс: в виде слияния клеток — копуляция — ил» обмен наследственным материалом — конъюгация.
Большинство простейших имеют одно ядро, но встречаются и многоядерные формы. Ядра некоторых простейших характеризуются полиплоидностью.
В жизненном цикле большинства простейших выделяют стадию трофозоита — активно питающуюся и перемещающуюся форму — и стадию цисты. Циста — неподвижная форма жизненного цикла простейших, покрытая плотной оболочкой и характеризующаяся резко замедленным обменом веществ. Паразитические простейшие инцисти-руются, попадая во внешнюю среду. В таком состоянии они способны переноситься ветром, водой и животными на огромные расстояния и таким образом расселяться. При попадании цисты в благоприятные условия происходит эксцистирование и простейшее начинает активно функционировать в состоянии трофозоита.
В настоящее время известно около 10 000 видов простейших. Основными средами их обитания являются вода и почва. Многие простейшие перешли к паразитическому или к комменсальному образу жизни.
Болезни, вызываемые простейшими, называют протозойными. Большинство простейших имеют время генерации от 6 до 24 ч. В связи с этим их размножение в организме хозяина обычно сопровождается экспоненциальным увеличением размеров их популяций до тех пор, пока этот процесс не замедлится или не остановится защитными механизмами хозяина или другими внешними факторами. Это означает, что один паразитический организм в принципе способен, размножившись, привести к гибели своего хозяина. В этом плане простейшие — возбудители заболеваний — сходны с возбудителями инфекционных болезней, например с патогенными бактериями и вирусами.
Медицинское значение имеют простейшие, относящиеся к классам Саркодовые, Жгутиковые, Инфузории и Споровики.
Один из разделов медицинской паразитологии – медицинская протозоология. Она изучает паразитических простейших, вопросы патогенеза, терапии заболеваний, диагностики.
Простейшие обитают повсеместно, где есть влажная среда.
Простейшие – животные организмы, имеющие микроскопические размеры, состоящие из 1 клетки. Простейшие имеют строение, типичное для эукариотической клетки. Есть протоплазма (эктоплазма – гомогенный наружный слой, эндоплазма – зернистый, с включениями, внутренний слой). На поверхности эктоплазмы образуется тонкая кожица – пелликула – гибкая структура, полупроницаемая, тонкая. Она позволяет временно изменять форму клетки. У некоторых есть кутикула, выполняющая защитную, структурную и опорную функции.
Функционально простейшие – сложные организмы, которые непрерывно взаимодействуют с окружающей средой. Они существенно отличаются от клеток многоклеточных организмов. Функции клеток определяются белками, которые синтезируются клетками. Клетка многоклеточного организма выполняет только определенные функции, поэтому в ней синтезируется только определенная часть белков. Клетка простейшего синтезирует все белки и ферменты, свойственные организму, в течение жизни работает весь ее геном, а не часть, как в клетке многоклеточного организма.
Простейшим присущи все свойства живых организмов. По тип обмена веществ и способу питания выделяют 2 группы:
– автотрофы;
– гетеротрофы.
Паразитические простейшие, как правило, питаются осмотически, поглощая вещества всей поверхностью. Переваривание осуществляется в пищеварительных вакуолях (пищевая частица набор ферментов в эндоплазме).
Сократительная вакуоль регулирует осмотическое давление клетки, это органелла выделения.
Простейшим присуща раздражимость. Они реагируют на механические, термические, световые, химические и другие воздействия. Часто реакция выражается в изменении направления движения – таксис. Если движение осуществляется в направлении действия раздражителя – положительный таксис, если наоборот – отрицательный. Таксисы позволяют простейшему перемещаться в свойственное место обитания, что чрезвычайно важно для паразита. Таксис определяет питание определенными веществами.
Для простейших характерно половое и бесполое размножение. В основе обоих процессов лежит клеточное деление, у многих видов простейших чередуются формы размножения, образуются сложные циклы.
Большинство простейших передвигаются. Имеются:
– постоянные органоиды движения – жгутики, реснички;
– временные – ложноножки – временные выросты эндоплазмы.
При неблагоприятных условиях простейшее перестает питаться, у него исчезают органоиды движения, образуется толстая оболочка – циста. Сам процесс носит название – инцистирование и позволяет переносить неблагоприятные условия, способствует расселению.
Простейшие имеют много общих черт, но отличаются по разным признакам (устройство органелл движения, тип питания и др.).
Тип Простейшие делится на 4 класса: 25000-35000
Sarcodina = Rhisopoda
Flagellata = Mastigophora
Sporozoa
Ciliala = Infusoria
Класс Саркодовые Sarcodina
Представители этого класса — самые примитивные простейшие. Форма их тела непостоянна. Передвигаются они с помощью ложноножек. Обитают в пресных водах, в почве, морях. В биогеоценозах выполняют функции консументов и редуцентов. Некоторые саркодовые адаптировались к комменсальному и паразитическому образу жизни. Медицинское значение имеют представители отряда амеб Amoebina. Паразитические амебы обитают у человека в основном в пищеварительной системе. Некоторые саркодовые, ведущие свободный образ жизни и обитающие в почве и загрязненной воде, при попадании в организм человека могут вызывать тяжелые заболевания, нередко заканчивающиеся смертью.
Класс Жгутиковые Flagellata
Тело жгутиковых кроме цитоплазматической мембраны покрыто еще и пелликулой — специальной оболочкой, обеспечивающей постоянство его формы. Имеется один или несколько жгутиков, органелл движения, представляющих собой нитевидные выросты эктоплазмы. Внутри жгутиков проходят фибриллы из сократительных белков. Некоторые жгутиковые имеют также ундулирующую мембрану — своеобразную органеллу передвижения, в основе которой лежит тот же жгутик, не выступающий свободно за пределы клетки, а проходящий по наружному краю длинного уплощенного выроста цитоплазмы. Жгутик приводит ундулирующую мембрану в волнообразное движение. Основание жгутика всегда связано с кинетосомой, органеллой, выполняющей энергетические функции. Ряд жгутиковых имеет также и опорную органеллу — аксостиль — в виде плотного тяжа, проходящего внутри клетки.
Разные виды паразитических жгутиковых у человека обитают в различных органах. Циклы их развития очень разнообразны.
Класс Инфузории Infusoria
Для инфузорий, как и для жгутиковых, характерно наличие пелликулы, им свойственна постоянная форма тела. Органеллы передвижения — многочисленные реснички, покрывающие все тело и представляющие собой полимеризованные жгутики. У инфузорий обычно два ядра: крупное — макронуклеус, регулирующее обмен веществ, и малое — микронуклеус, служащее для обмена наследственной информацией при конъюгации. Макронуклеусы инфузорий полиплоидны, микронуклеусы — гаплоидны или диплоидны. Сложно организован аппарат пищеварения. Имеется постоянное образование: клеточный рот — цито-стом, клеточная глотка — цитофаринкс. Пищеварительные вакуоли перемещаются по эндоплазме, при этом литиче”кие ферменты выделяются поэтапно. Это обеспечивает полноценное переваривание пищевых частиц. Непереваренные остатки пищи выбрасываются через порошицу — специализированный участок клеточной поверхности.
Инфузории — наиболее высоко организованные простейшие. Паразитов среди них относительно немного. У человека паразитирует единственная инфузория — балантидий, которая обитает в пищеварительной системе.
Класс Споровики Sporozoa
Все споровики — паразиты и комменсалы животных и человека. Органеллы движения у них отсутствуют. Питание споровиков осуществляется за счет поглощения пищи всей поверхностью тела. Многие споровики — внутриклеточные паразиты. Они претерпели наиболее глубокую дегенерацию. Цикл развития включает стадии бесполого размножения, нолового процесса в виде копуляции и спорогонии. Бесполое размножение осуществляется путем простого или множественного деления — шизогонии. Половому процессу предшествует образование половых клеток — мужских и женских гамет. Гаметы сливаются, а образовавшаяся зигота покрывается оболочкой, под которой происходит спорогония — множественное деление с образованием спорозоитов (рис. 19.1).
Ниже описаны паразитические и комменсальные простейшие, обитающие в разных органах человека. От специфики органа, являющегося средой обитания паразита, зависят пути проникновения и патогенное действие паразита, методы диагностики соответствующих заболеваний и меры их профилактики. Поэтому простейшие, с медицинской точки зрения, могут быть разделены на виды, обитающие в полостных органах, которые имеют связь с внешней средой, и живущие в тканях внутренней среды человека. Кроме того, выделяют группу свободноживущих простейших, случайное попадание которых в организм человека может приводить к острейшим патологическим процессам и даже к смерти. Соответствующие три экологические группы простейших описаны отдельно.
130.Амеба дизентерийная, Неглерия, Акантамёба. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Sarcodina
Entamoeba histolylica — возбудитель амебиаза. Амебиаз встречается повсеместно, но чаще в зонах с влажным жарким климатом. В цикле развития амебы имеется несколько стадий, морфологически и физиологически отличающихся друг от друга. Мелкая вегетативная форма обитает в просвете кишки. Размеры ее 8—20 мкм. В цитоплазме можно обнаружить бактерии и грибки — элементы микрофлоры кишечника.
Крупная вегетативная форма также обитает в просвете кишки в гнойном содержимом язв кишечной стенки. Ее размеры — до 45 мкм. Цитоплазма четко разделена на прозрачную, стекловидную эктоплазму и зернистую эндоплазму. В ней расположены ядро с характерной темно окрашенной кариосомой и эритроциты, которыми она питается. Крупная форма энергично передвигается с помощью широких псевдоподий. В глубине пораженных тканей располагается тканевая форма. Она мельче крупной вегетативной формы и не имеет в цитоплазме эритроцитов. Цисты обнаруживаются в фекалиях хронически больных и паразитоносителей, у которых заболевание проходит бессимптомно. Цисты имеют округлую форму диаметром 8—15 мкм и от одного до четырех ядер в виде колечек.
Жизненный цикл паразита сложен (рис. 19.4). Человек заражается амебиазом, проглатывая цисты паразита с водой или пищевыми продуктами, загрязненными землей. В просвете толстой кишки из цисты образуется, за счет следующих друг за другом делений, восемь мелких клеток, превращающихся в мелкие вегетативные формы. Вреда человеку они не приносят. Они могут вновь инцистироваться и выходить наружу. При ухудшении условий существования хозяина мелкие вегетативные формы способны превращаться в крупные, которые вызывают образование язв. Погружаясь глубже, они превращаются в тканевые формы, которые в особо тяжелых случаях могут попадать в кровь и разноситься по всему организму. При этом возможно образование абсцессов в печени, легких и других органах. В остром периоде заболевания у больного в фекалиях обнаруживаются не только цисты, но и трофозоиты.
Диагноз ставится на основе обнаружения в фекалиях трофозоитов с заглоченньми эритроцитами. Четырехъядерные цисты могут свидетельствовать скорее о хроническом течении заболевания или о пара-зитоносительстве.
Профилактика — как при лямблиозе.
Дизентерийная амеба. /Entamoeba histolitica/
– открыта русским ученым в Петербургской военной академии. Заболевание – амебиаз, представляющее язвенное поражение толстой кишки. В запущенных случаях возможно прободение кишки. Осложнения – перитонит и абсцессы печени.
Жизненный цикл амебы включает вегетативную стадию и цисту. Стадии переходят друг в друга в зависимости от условий. Вегетативная форма включает следующие формы:
– тканевая;
– большая вегетативная (forma magna);
– малая вегетативная (forma minuta);
– предцистная.
Тканевая форма – 20-25 мкм. В центре – звездчатая кариосома. Движения в виде толчков. Обнаруживается при остром амебиазе в тканях стенки кишечника. Эритроциты внутри клетки присутствуют.
Большая вегетативная – 30-40 мкм. Тело четко делится на экто- и эндоплазму, ядро практически не видно. Всегда обнаруживаются фагоцитированные эритроциты от 1 до 20 штук. Поэтому – эритрофаг/ гематофаг. Определяются при остром амебиазе из свежевыделенных фекалий больного человека.
Мелкая вегетативная форма. Живет в просвете кишечника, питается бактериями, структуры хозяина в ней не обнаруживаются, следовательно, эритроцитами не питается. Отношения с хозяином – носительство. Это комменсальная форма, размеры которой около 13 мкм.
Предцистная – 13-15 мкм. Протоплазма гомогенизирована, свободна от каких-либо включений.
Вне организма хозяина все формы быстро погибают.
Стадия покоя – циста 2-5 мкм. Цисты – незрелые 1-2 ядерные, зрелые – 4-ядерные (инвазионная стадия для хозяина). Стадии цисты очень устойчивы. Цисты образуются в нижних отделах толстого кишечника.
Заражение человека осуществляется цистами. Инкубационный период составляет от недели до 3 месяцев. Из цисты выходят мелкие просветные формы. Микротравмы, воспаления слизистой, интоксикация, травмы, ранения, нарушения водного и пищевого обмена способствуют переходу просветной формы в тканевую. Под действием лизирующих ферментов разрушается стенка кишечника, в просвет начинает выделяться кровь. Возникновение крупной вегетативной формы – не причина амебиаза. Появление её – результат далеко зашедшего некроза кишечной стенки.
Диагноз ставится на основании обнаружения в фекалиях вегетативных форм (обеих) и цист различной степени зрелости.
– Повсеместно
– попадание цист в пищеварительный тракт
– локализация: кишечник
– обнаружение 4х ядерных цист и вегетативных в фекалиях
– выделение и лечение цистовыделителей и цистоносителей, сангигиен культура населения
131.Комменсальные и условно-патогенные простейшие: Амеба кишечная, Амеба ротовая.
тип Protozoa
класс Sarcodina
§
Размер около 20 мкм. Эндоплазма грубо вакуолизирована. В вакуолях – бактерии, грибы, простейшие других видов. Зрелые цисты имеют 8 ядер, незрелые – 2. иногда встречаются формы с 16-32 ядрами. Комменсал.
Ротовая амеба/Entamoeba gingivalis
Часто встречается в кариозных полостях зубов и в белом мягком налете. Похожа на дизентерийную. Питается бактериями, эритроцитами, лейкоцитами. Вопрос о патогенности не решен.
Ротовая амеба Entamoeba gingivalis (кл. Саркодовые) — комменсал, обитающий на деснах, зубном налете и в криптах нёбных миндалин более чем у 25% здоровых людей (рис. 19.2, А). У лиц с заболеванием полости рта встречается чаще. Размеры клетки 6—30 мкм, псевдоподии широкие. Питается бактериями и лейкоцитами, при кровотечении из десен может захватывать и эритроциты. Цист не образует.
132.Трихомонады. Систематика, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Flagellata
Trichomonas hominis
Возбудитель кишечного трихомониаза. Размер 5-15 мкм. Форма тела овально- уплощенная. Есть жгутики, ундулирующая мембрана. Питается бактериями, эритроцитами, обитает в толстом кишечнике.
Trichomonas vaginalis
Возбудитель урогенитального трихомониаза. Размер 15-20 мкм. Локализуется в мочеполовой системе мужчин и женщин. Зараженность женщин составляет 20-40%, мужчин – до 15%. Чаще всего заражаются в 18-45 лет. У женщин наблюдаются патологические явления во влагалище, у мужчин – повреждается предстательная железа, что ведет к бесплодию. Передается половым путем, а также при нестерильном гинекологическом осмотре.
Обнаружение происходит по мазку из выделений.
В половых органах человека обитает влагалищная трихомонада Trichomonas vaginalis (кл. Жгутиковые) — возбудитель трихомоноза. Длина этого паразита 14—30 мкм. Форма тела грушевидная. На переднем конце находятся четыре жгутика. До середины клетки доходит также небольшая ундулирующая мембрана. По середине тела тянется аксостиль, выступающий из клетки на ее заднем конце. Характерна форма ядра, овального, заостренного с двух концов, и напоминающего косточку сливы (рис. 19.5).
В пищеварительных вакуолях располагаются лейкоциты, эритроциты и бактерии, которыми этот паразит питается. Цист не образует. Эта трихомонада обитает у женщин во влагалище и в шейке матки, а у мужчин — в мочеиспускательном канале, мочевом пузыре и в предстательной железе. Зараженность женщин достигает 20—40%, мужчин — 15%. Серьезных повреждений хозяину эта трихомонада не наносит, но, тесно контактируя с эпителием мочеполовой системы, она вызывает врзникновение мелких воспалительных очагов под эпителиальным слоем и слущивание поверхностных клеток слизистой оболочки. Через нарушенную эпителиальную выстилку в просвет органа поступают лейкоциты. У мужчин заболевание обычно завершается спонтанным выздоровлением примерно через 1 мес. У женщин трихомоноз может протекать несколько лет.
Лабораторная диагностика — обнаружение живых подвижных трихомонад в мазке из выделений мочеполовых путей.
Профилактика — соблюдение правил личной гигиены при половых контактах.
– Повсеместно
– при половом общении
– локализация: МПС м и ж
– обнаружение вегетативных форм в выделениях и соскобах из мочеполовых путей
– личная и общественная гигиена
133.Трипаносомы. Систематика, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики
тип Protozoa
класс Flagellata
Гамбийская трипаносома.
Возбудитель африканского трипаносомоза – смертельного заболевания Тропической Африки. Основной источник – человек, носители – антилопы, свиньи, переносчик – муха цеце. Трипаносомы имеют размеры 20-30 мкм. Есть удлиненное тело, ундулирующая мембрана, свободный ее конец. Когда муха цеце сосет кровь больного, происходит попадание в ее желудок трипаносом. Он6и размножаются и сложно изменяются – инвазионная для млекопитающих форма. При укусе попадает зараженная слюна и трипаносомы проникают в кровь человека. Чаще всего они оседают в головном мозге, поражается ЦНС, нарастает сонливость, происходят патологические изменения в печени, почках, легких и других органах. Болезнь длится 5 – 7 лет и без лечения заканчивается летально. Диагностика болезни по пунктатам спинного мозга, лимфатических узлов и периферической крови.
Trypanosoma brucei gambiense и T.b. rhodesiense (кл. Жгутиковые) — возбудители африканского трипаносомоза, или сонной болезни. Паразит имеет извилистую заостренную с обеих сторон форму. Длина его 17—28 мкм. Стадии, паразитирующие у человека, имеют один жгутик, ундулирующую мембрану сбоку и хорошо заметный кинетопласт у основания жгутика.
Трипаносомы поселяются у человека в крови, лимфе, спинномозговой жидкости, в тканях головного и спинного мозга и в серозных полостях. T.b. gambiense встречается в Западной Африке, а T.b. rhodesiense — в Восточной и Юго-Восточной Африке.
Жизненный цикл этих паразитов протекает в организме человека, домашних и диких млекопитающих, в первую очередь копытных. T.b. gambiense чаще поражает человека, свиней и собак, T.b. rhodesiense — диких животных — антилоп и носорогов. Переносчиком первого подвида является муха це-це ,Glossina palpalis, живущая поблизости от жилища человека, второго— G. morsitans, обитающая в открытых саваннах и саванновых лесах. В связи с этим сонная болезнь, возбудителем которой является T.b. gambiense, встречается в антропогенных очагах культурных ландшафтов. Ежегодно регистрируется около 10 000 новых случаев заражения. Восточноафриканский трипаносомоз распространен значительно реже в естественной природе. В основном заболевают охотники, туристы, сезонные рабочие, каждый год — около 1500 человек.
Сонная болезнь без лечения протекает около 5 лет и выражается в нарастающей мышечной слабости, депрессии, истощении и сонливости. Возможны случаи самоизлечения, но обычно заболевание заканчивается смертью больного.
Восточноафриканский трипаносомоз протекает более злокачественно, длится не более 6 мес. и также заканчивается смертью.
Для паразитирования трипаносом у млекопитающих и человека характерны циклические подъемы интенсивности инвазии за счет их размножения, сопровождающиеся изменениями строения и антигенных свойств паразитов. Во время увеличения количества паразитов в крови преобладают трипаносомы удлиненной формы. Антигены, которые они образуют, вызывают формирование антител в организме хозяина. Под действием антител многие паразиты гибнут и интенсивность инвазии снижается. Выжившие трипаносомы укорачиваются и начинают вырабатывать другие антигены. Укороченные формы паразита, инвазионные для мухи це-це, в ее организме вновь приобретают удлиненную форму, инвазионную для человека. Изменение формы тела и смена антигенных свойств оболочки повторяются многократно. Таким образом, популяция паразита в хозяине выживает и избегает его иммунной реакции.
Антигенные свойства поверхности трипаносомы зависят только от одного белка — гликопротеина, полностью покрывающего всю клетку. Гликопротеин построен из 470 остатков аминокислот. Каждая новая волна размножения паразитов представляет собой новую популяцию трипаносом, обладающих новым поверхностным антигеном. Эти вариации антигенных свойств помогают паразиту преодолевать иммунный ответ хозяина и делают невозможной вакцинацию населения, обитающего в природных очагах трипаносомозов.
Смена антигенных свойств обеспечивается заменой поверхностных гликопротеинов, кодируемых разными генами, относящимися к одному мультигенному семейству. Один клон трипаносом может образовывать попеременно до 100 разных варьирующих гликопротеинов. Гены этих протеинов возникли в процессе эволюции, вероятно, путем дупликаций и последующей дифференцировки, как и другие семейства генов. В геноме трипаносомы имеется сайт экспрессии, в который поочередно перемещаются гены поверхностных гликопротеинов, приближаясь к промотору, обеспечивающему их специфическую активацию. Там они транслируются. Не исключено, однако, что сайт экспрессии в геноме трипаносомы не единственный, и даже возможно, что разные гены гликопротеинов активируются несколькими механизмами. В любом случае речь идет о своеобразной адаптации паразита к специфическим условиям существования, повышающей его выживаемость и открывающей ему широкие эволюционные перспективы.
Лабораторная диагностика — исследование мазков крови и спинномозговой жидкости больного для выявления в них возбудителя. Используются также иммунологические реакции и заражение лабораторных животных.
Профилактика — кроме борьбы с переносчиками применяют профилактическое лечение здоровых людей, живущих в очагах трипаносомоза, делающее организм невосприимчивым к инвазии. Практиковавшийся ранее отстрел диких животных, являющихся природным резервуаром паразита, вряд ли рационален в связи с возможным нарушением экологического баланса, который складывался в биогеоценозах на протяжении тысячелетий.
Trypanosoma cruy — возбудитель американского трипаносомоза, или болезни Чагаса. Длина этой трипаносомы в крови человека достигает 20 мкм. Кинетопласт очень крупный, округлой формы. Характерной особенностью возбудителя является способность к внутриклеточному паразитизму. При этом трипаносомы проникают вначале в макрофаги кожи и слизистых оболочек, а затем и в клетки миокарда, нейроглии и мышц, теряя жгутики, ундулирующие мембраны, и превращаясь в безжгутиковые, или амастиготные, формы. Здесь и происходит размножение паразитов. В крови эти трипаносомы никогда не делятся. В конечном счете, пораженная клетка вся заполняется амастиготными формами трипаносом и разрывается, а паразиты инвазируют новые клетки. При этом часть их, превращаясь вновь в жгутиковую форму, поступает в кровь, откуда в дальнейшем они могут попасть в организм переносчика.
134.Лямблия кишечная. Систематика, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Flagellata
Lamblia intestinalis
Открыта профессором Харьковского университета Лямблем в 1859 году. Мелкие жгутиконосцы – 10-20 мкм. Локализуются в ДПК. Имеет грушевидную форму, 2 ядра, все органеллы парные. На вентральной стороне имеется присасывательный диск. Облигатные паразиты. Заражение происходит при попадании цист. Лямблиоз широко распространен, особенно у детей. Зараженность достигает 50-80%. Интенсивность инвазии очень велика (на 1 кв. см. 1000000 особей).
Диагноз ставится по обнаружению вегетативных форм и цист в фекалиях и при дуоденальном зондировании.
В тонкой кишке человека паразитирует единственный вид простейших — лямблия Lamblia intestinalis (кл. Жгутиковые) — возбудитель лямблиоза, которым чаще болеют дети. Форма паразита напоминает грушу, разрезанную вдоль. Длина тела 10—18 мкм. В расширенной части, на уплощенной стороне расположен присасывательный диск, с помощью которого лямблии присасываются к ворсинкам кишечника. Вдоль тела проходят две тонкие опорные органеллы — аксостили. Симметрично в клетке располагаются два ядра и четыре пары жгутиков (рис. 19.3, А).
Трофозоиты используют питательные вещества с поверхности клеток кишечного эпителия. Захват пищи осуществляется пиноцитозом.
Большие количества лямблии, которые покрывают обширные поверхности кишечной стенки, нарушают процессы всасывания и присте-ночного пищеварения. Попадая в нижние отделы тонкой кишки, лямблии инцистируются. Зрелые цисты имеют овальную форму, четыре ядра и несколько аксостилей. Во внешней среде цисты сохраняют жизнеспособность в течение нескольких недель. Заражение человека происходит при проглатывании цист.
Лабораторная диагностика — обнаружение цист в фекалиях и тро-фозоитов в содержимом двенадцатиперстной кишки, полученном при дуоденальном зондировании.
Личная профилактика — соблюдение правил гигиены питания. Общественная профилактика — санитарное благоустройство туалетов, предприятий общественного питания.
– Повсеместно
– цистами при попадении их на продукты или питьевой водой
– локализация: 12 перстная
– обнаружение вегетативных форм в фекалиях и 12перстной кишке при зондировании
– личная гигиена
135.Лейшмании. Систематика, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Flagellata
Паразиты рода Лейшмания.
Обитают в организме позвоночных, где они встречаются в лейшманиальной безжгутиковой внутриклеточной форме. В их организме превращаются в лептомональную жгутиковую форму. Попадая в организм человека, вызывают лейшманиозы – трансмиссивные тропические и субтропические заболевания человека и животных.
Лейшмания доновани, 1903г. Селезенка больного человека. Индийский кала-азар, черная лихорадка, дум-дум, висцеральный лейшманиоз.
Попадая в организм с кровью больного, лейшмании образуют жгутиковую форму.
Висцеральный лейшманиоз часто поражает детей до 12 лет. У переболевших вырабатывается стойкий иммунитет. Течение болезни характеризуется анемией, лейкопенией, волнообразной лихорадкой. Встречается в Индии, Кении, Азии, Казахстане Закавказье.
Диагностика – пунктаты костного мозга, лимфатических узлов, мазки крови.
Тропическая лейшмания.
Болезнь Боровского (1898), кожный лейшманиоз.
Характеризуется поражениями кож, образованием язв, после которых остаются рубцы. Размер язв до 10-15 см. существует антропонозный, поздно проявляющийся подтип кожного лейшманиоза, встречающийся в городах и поселках городского типа на Ближнем Востоке, Южной Европе, индии. Основной источник – человек, переносчик – москиты. Язвы образуются на открытых участках тела, затем они рубцуются. Зоонозный подтип (раноизъязвляющийся, полупустынно-сельский, остро некротизирующий, Пендинская язва). Имеет сезонность с лётом москитов. Резервуар – мыши, суслики, особенно – большая песчанка. Москиты питаются кровью в сумерки, в природе существует очаг. При зоонозном подвиде чаще поражаются нижние конечности. Язвы могут также покрывать все тело. Подтип распространен в Азии, Африке, на юге России, Туркмении, Узбекистане.
Диагностика – по обнаружению простейших в кожных пробах вокруг ран.
Лейшмании Leischmania (кл. Жгутиковые) — возбудители лейшманиозов. Заболевания человека вызываются несколькими видами и подвидами паразитов, которые объединяются в четыре комплекса: L. donovani — возбудитель висцерального лейшманиоза, L. tropica — возбудитель кожного лейшманиоза, L. mexicana — возбудитель лейшманиоза Центральной Америки, L. brasiliensis — возбудитель бразильского лейшманиоза. Все виды сходны морфологически и имеют одинаковые циклы развития. Они существуют в двух формах: в безжгутиковой, или лейшманиальной, и жгутиковой; или промастиготной (рис. 19.8).
Лейшманиальная форма очень мелка — 3—5 мкм в диаметре. Характерной чертой ее является круглое ядро, занимающее около ‘/4 цитоплазмы; жгутика нет, но перпендикулярно клеточной поверхности располагается палочковидный кинетопласт. Эти формы обитают в клетках ретикулоэндотелиальной системы человека и ряда млекопитающих (грызунов, собак, лис). Промастиготная форма удлинена — до 25 мкм, спереди находится жгутик, у основания которого хорошо виден такой же кинетопласт, что и в безжгутиковой стадии паразита. Обитает в пищеварительной системе москитов. Безжгутиковая форма, посеянная на культуральную среду, превращается в жгутиковую.
Лейшманиозы широко распространены в странах с тропическим и субтропическим климатом на всех континентах там, где обитают москиты. Они—типичные природно-очаговые заболевания (см. § 18.13). Природными резервуарами являются грызуны, дикие и домашние хищники. Заражение человека происходит при укусе инвазированными москитами.
По патогенному действию лейшманий заболевания, которые они вызывают, делят на три основные формы: кожный, слизисто-кожный и висцеральный лейшманиозы.
При кожном лейшманиозе очаги поражения находятся в коже. Это самый распространенный тип лейшманиоза, протекающий относительно доброкачественно. Возбудителями кожного лейшманиоза в Африке и Азии являются L. tropica, а в Западном полушарии — L. mexicana и ряд штаммов L. brasiliensis. Лейшманий L. tropica и L. mexicana вызывают на коже длительно не заживающие язвы на месте укусов москитами. Язвы заживают через несколько месяцев после образования, а на их месте на коже остаются глубокие рубцы. Некоторые формы L. brasiliensis способны распространяться по лимфатическим сосудам кожи с образованием многочисленных кожных язв в отдалении от мест укусов.
Слизисто-кожный лейшманиоз вызывается подвидом L. brasiliensis brasiliensis. При этой форме заболевания паразиты проникают из кожи по кровеносным сосудам в носоглотку, гортань, мягкое нёбо, половые органы, поселяются в макрофагах соединительных тканей этих органов и вызывают здесь деструктивные воспаления.
Висцеральный лейшманиоз вызывает L. donovani. Заболевание начинается через несколько месяцев или даже лет после заражения как системная инфекция. Паразиты размножаются в макрофагах и в моноцитах крови. Нарушаются функции печени, кроветворение. Очень велика интоксикация. При отсутствии лечения заболевание заканчивается смертью.
Лабораторная диагностика основана на микроскопировании мазков из кожных язв при кожном и слизисто-кожном лейшманиозах, пунктатов лимфатических узлов и костного мозга при висцеральном лейшманиозе. В окрашенных препаратах обнаруживается лейшманиальная форма паразитов как внутри клеток, так и внеклеточно. В сомнительных случаях производят посев материала, взятого от больного, на специальную культуральную среду, на которой лейшманий приобретают промастиготную форму, активно передвигаются и легко обнаруживаются при микроскопировании. Используют также и биологические пробы — заражение лабораторных грызунов.
Профилактика — в первую очередь, это борьба с переносчиками и уничтожение природных резервуаров (грызунов и бродячих собак), а также профилактические прививки.
Доновани:
– индия, средиземноморье, закавказье, ср. азия
– при укусе москита
– локализация: клетки внутренних органов (печень, селезенка, км)
– микроскопирование мазков пункций грудины и лимфоузлов
– защита от москитов
Тропика:
– европа, азия, америка с субтропическим климатом
– при укусе москита
– локализация: клетки кожи
– микроскопирование мазков из отделяемого язв
– защита от москитов
136.Малярийные плазмодии. Систематическое положение, морфология, географическое распространение, цикл развития, видовые отличия. Борьба с малярией. Задачи противомалярийной службы на современном этапе.
тип Protozoa
класс Sporozoa
Малярийные плазмодии Plasmodium (кл. Споровики) — возбудители малярии. Известны следующие виды малярийных плазмодиев, паразитирующие у человека: Р. vivax — возбудитель трехдневной малярии, Р. falciparum — возбудитель тропической малярии, Р. malariae — возбудитель четырехдневной малярии, Р. ovale— возбудитель овале-малярии, близкой к трехдневной. Три первых вида широко распространены в тропических и субтропических климатических поясах, последний — только в тропической Африке. Все виды сходны морфологически и жизненными циклами, отличаясь друг от друга деталями строения и некоторыми особенностями цикла развития, проявляющимися в основном продолжительностью его отдельных периодов.
Жизненный цикл малярийных плазмодиев типичен для споровиков, включая стадии бесполого размножения в виде шизогонии, полового процесса и спорогонии. Окончательным хозяином паразитов является комар р. Anopheles (см. разд. 21.22), а промежуточным — только человек. Комар является одновременно и переносчиком. Поэтому малярия — типичное антропонозное трансмиссивное заболевание.
Со слюной зараженного комара при укусе плазмодии попадают в кровь человека (рис. 19.10). Развитие паразитов в организме человека происходит синхронно. С током крови они разносятся по организму и поселяются в клетках печени. Здесь они растут и размножаются шизогонией таким образом, что один паразит делится на тысячи дочерних особей. Клетки печени при этом разрушаются и паразиты, называющиеся на этой стадии мерозоитами, поступают в кровь и внедряются в эритроциты. С этого момента начинается эритроцитарная часть цикла развития плазмодия. Паразит питается гемоглобином, растет и размножается шизогонией. При этом каждый плазмодий делится на 8—24 мерозоита. После разрушения эритроцита мерозоиты попадают в плазму крови и оттуда в новые эритроциты, после чего весь цикл эритроцитарной шизогонии повторяется.
Рис. 19.10. Жизненный цикл малярийного плазмодия:
1 — преэритроцитарная шизогония в клетках печени, 2 — эритроцитарная шизогония, 3 — образование гаметоцитов, 4 — оплодотворение, 5 — спорогония в стенке желудка комара, 6 — овоциста со споромитами, 7 — проникновение спорозоита в слюнные железы комара, 8— заражение человека
Из части мерозоитов в эритроцитах образуются незрелые половые клетки — мужские и женские гаметоциты. Они являются инвазионной стадией для комара. Дальнейшее их развитие возможно только в его пищеварительной системе. При укусе больного человека комаром гаметоциты попадают в желудок последнего, где из них образуются зрелые гаметы. В результате оплодотворения в желудке комара образуется подвижная зигота, которая перемещается на наружную поверхность стенки желудка и покрывается оболочкой, формируя ооцисту. С этого момента начинается период спорогонии, когда содержимое ооцисты многократно делится, образуя около 10000 спорозоитов — тонких серповидных клеток, которые после разрыва оболочки поступают в слюнные железы комара. При кровососании спорозоиты поступают в кровяное русло человека.
Таким образом, в организме человека плазмодий размножается только бесполым путем — шизогонией, человек является его промежуточным хозяином. В организме комара проходят две другие стадии цикла развития паразита: половой процесс — гаметогония и образование спорозоитов за счет деления под оболочкой ооцисты — спорогония. Поэтому малярийный комар является окончательным хозяином этого паразита.
Выход большого количества мерозоитов из эритроцитов сопровождается выбросом в плазму крови значительной массы токсических продуктов жизнедеятельности. Их воздействие на организм приводит к резкому повышению температуры, ознобу, слабости и головным болям. Такое состояние возникает внезапно и длится в среднем 1,5—2 ч. Вслед за этим наступает чувство жара, сухость во рту, жажда. Температура тела достигает 40—41°С. Через несколько часов все перечисленные симптомы исчезают, и больные обычно засыпают. Весь приступ может продолжаться от 6 до 12 ч. При трехдневной и овале-малярии промежутки между приступами составляют 48 ч, число таких приступов может достигать 10—15, после чего они прекращаются за счет повышения уровня специфического иммунитета, но паразиты в крови еще могут обнаруживаться. В таком случае человек становится паразитоносителем и продолжает представлять опасность для окружающих как возможный источник заражения.
Естественный отбор приводит к возникновению новых антигенных вариантов возбудителя, которые обеспечивают возможность наступления рецидивов заболевания. Рецидивы могут повторяться несколько раз, но постепенно популяция эритроцитарных паразитов полностью погибает. Однако в течение 3—5 лет инвазия может вновь активизироваться за счет находящихся в латентном состоянии в печени экзоэритроцитарных шизонтов, которые могут выходить из печеночных клеток и внедряться в эритроциты. Таким образом, весь процесс болезни может начаться снова.
При малярии, вызываемой Р. malariae, приступы повторяются через 72 ч. Часто встречается и бессимптомное носительство. Экзоэритро-цитарной стадии в цикле развития этого паразита нет, поэтому поздние рецидивы невозможны, хотя инвазия характеризуется упорным течением и длится до 40 лет.
При тропической малярии вначале приступы развиваются через разные промежутки времени, а позже — через 24 ч. От осложнений со стороны центральной нервной системы или почек возможна смерть больного. Шизонты в клетках печени не сохраняются, а заболевание может продолжаться до 18 мес.
Все виды малярийных плазмодиев могут инвазировать человека и при гемотрансфузии (переливание крови). В этом случае ни у одного из паразитов не формируется экзоэритроцитарной стадии. Поэтому поздних рецидивов в этом случае не бывает. Гемотрансфузионный способ заражения наиболее часто встречается при четырехдневной малярии в связи с тем, что при этой форме болезни шизонты в эритроцитах находятся в очень малом количестве и могут не обнаруживаться при исследовании крови доноров.
Иногда человек может быть инвазирован одновременно двумя или тремя видами плазмодиев. В таком случае малярийные приступы не имеют четкой периодичности и клинический диагноз затруднен.
Лабораторный диагноз малярии можно поставить только в период, соответствующий стадии эритроцитарной шизогонии, когда в крови удается обнаружить паразитов.
Плазмодий, недавно проникший в эритроцит, имеет кольцевидную форму. Его цитоплазма выглядит как ободок, окружающий крупную вакуоль с продуктами диссимиляции. Ядро паразита смещено к краю клетки. Следующая стадия называется амебовидным шизонтом. У паразита появляются ложноножки, а вакуоль увеличивается. Наконец плазмодий занимает почти весь эритроцит. Следующая стадия развития паразита — фрагментация шизонта. На фоне деформированного эритроцита обнаруживаются множественные мерозоиты, в каждом из которых лежит ядро. Кроме бесполых клеток в эритроцитах можно увидеть и гаметоциты. Они отличаются крупными размерами, не имеют псевдоподий и вакуолей (рис. 19.11).
Рис. 19.11. Малярийные плазмодии. Стадии развития в эритроцитах:
I—стадия кольца, II—стадия амебовидного шизонта, III—стадия фрагментации, IV—гаметоциты
Профилактика малярии — раннее выявление и лечение больных, профилактическое лечение в зонах широкого распространения малярии. Как и при любых трансмиссивных заболеваниях, необходима прицельная борьба с переносчиками.
137.Токсоплазма. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Sporozoa
Токсоплазма Toxoplasma gondii — возбудитель токсоплазмоза. Имеет форму полумесяца, один конец которого заострен более другого. В центре располагается крупное ядро. Длина паразита 4—7 мкм (рис. 19.7).

Рис. 19.7. Токсоплазма (Л) и саркоциста (Б):
1 — единичные паразиты, 2 — спорозоиты под общей оболочкой, 3 — изолированная спора, 4— саркоциста в мышечном волокне
Токсоплазма поражает огромное количество видов животных и человека. Иммунологические исследования показали, что на Земле токсоплазмами заражено более 500 млн. человек.
Жизненный цикл токсоплазмы типичен для споровиков: в нем чередуются стадии шизогонии, гаметогонии и спорогонии.
Основные хозяева паразита — домашние кошки и дикие виды сем Кошачьи. Они заражаются, поедая больных грызунов, птиц или инвазированное мясо крупных животных. Паразиты у них сосредоточиваются в клетках кишечника, размножаются шизогонией, а затем образуют гаметы. После копуляции гамет формируются ооцисты, которые выделяются во внешнюю среду. В них происходит спорогония, т. е. деление зиготы под оболочкой.
Такие спороцисты со спорозоитами рассеиваются кошками и попадают к промежуточным хозяевам, которыми могут быть человек, почти все млекопитающие, птицы и даже пресмыкающиеся. В клетках большинства их органов происходит бесполое размножение токсоплазм в форме множественного деления. В результате образуются группы, состоящие из многих сотен отдельных паразитов. Эти группы могут распадаться, и тогда отдельные токсоплазмы внедряются с помощью специфической органеллы проникновения — коноида — в непораженные клетки, в которых вновь происходит шизогония.
Другие такие группы покрываются плотной оболочкой и формируют цисты. Цисты очень устойчивы и могут длительное время находиться в состоянии покоя в органах хозяев. В окружающую среду они не выделяются. Цикл развития замыкается при поедании кошками органов промежуточных хозяев с цистами.
Своеобразной особенностью цикла развития токсоплазм является то, что промежуточные хозяева могут заражаться ими не только от основного хозяина, но и при поедани» друг друга. Так, возможно заражение свиней при поедании ими трупов грызунов, погибших от токсоплазмоза, грызуны же заражаются друг от друга при каннибализме. Возможно и внутриутробное заражение плода от больной беременной самки, когда паразиты проникают через плаценту. Этот способ заражения обеспечивает устойчивое существование природных очагов токсоплазмоза и среди мелких грызунов, не склонных к каннибализму.
В соответствии с этим и человек как промежуточный хозяин может заразиться токсоплазмозом разными путями: 1) при поедании мяса инвазированных животных; 2) с молоком и молочными продуктами; 3) через кожу и слизистые оболочки при уходе за больными животными, при обработке шкур и разделке животного сырья; 4) внутриутробно через плаценту; 5) при медицинских манипуляцих переливания крови и лейкоцитарной массы, при пересадках органов, сопровождающихся приемом иммунодепрессивных препаратов. Последнее свидетельствует о том, что общее снижение иммунитета повышает вероятность заражения токсоплазмозом.
Обычно паразиты обладают весьма низкой патогенностью, но в некоторых условиях они могут вызвать очень тяжелые нарушения, что зависит как от индивидуальной чувствительности хозяев, так и от путей проникновения токсоплазм в организм человека.
Наиболее опасным является трансплацентарное заражение. При этом возможно рождение детей с множественными врожденными пороками развития, в первую очередь головного мозга. При постановке диагноза используют методы иммунологических реакций, обнаружение токсоплазм при прямом микроскопировании материала, взятого от больного человека или трупа. Для исследования используют плаценту, печень, кровь, лимфатические узлы, головной мозг. Применяют также метод биологических проб. В этом случае лабораторным животным вводят кровь или спинномозговую жидкость больного. Мыши заболевают токсоплазмозом при таком способе заражения в острой форме, и обнаружение возбудителя у них не представляет сложности.
Профилактика — термическая обработка животных продуктов питания, санитарный контроль на бойнях и мясокомбинатах, предотвращение тесных контактов детей и беременных женщин с домашними животными.
Toxoplasma gandi
Возбудитель токсоплазмоза. Открыт в 1908 году. Проникает через кожу, слизистые, пищеварительный тракт. Локализуется практически во всех органах. Бывает врожденные формы. Кошачьи – окончательный хозяин, а человек – промежуточный. Смертность очень велика, выжившие люди становятся инвалидами. Все беременные исследуются на носительство. Зооноз с частой природной очаговостью.
138.Саркоцисты. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Protozoa
класс Sporozoa
Несколько близких к токсоплазме паразитов — саркоцисты (Sarcocystis hominis, S. suihominis, S. lindemanni) — являются возбудителями саркоцистозов и имеют сходный с ней цикл развития. Человек для этих паразитов является основным хозяином, а животные — промежуточными. Поэтому у человека, как и у кошки при токсоплазмозе, поражается кишечник. Но степень поражения его очень незначительна. По некоторым данным, частота инвазии людей саркоцистами достигает 7—60%, особенно там, где в соответствии с традициями население употребляет в пищу сырое или полусырое мясо. Врачи обычно не ставят правильного диагноза, а заболевания заканчиваются быстрым самоизлечением.
см. альбом!
139.Балантидий кишечный. Систематическое положение, цикл развития, географическое распространение, пути заражения, патогенное действие, методы лабораторной диагностики, меры профилактики.
тип Protozoa
класс Infusoria
Балантидий Balantidium coli (кл. Инфузории) — возбудитель балантидиаза. Это крупное простейшее, длиной до 200мкм. Сохранены многие признаки свободноживущих инфузорий: все тело покрыто ресничками, имеются цитостом и цитофаринкс. Под пелликулой расположен слой прозрачной эктоплазмы, глубже находится эндоплазма с органеллами и двумя ядрами. Макронуклеус имеет гантелевидную или бобовидную форму, рядом с ним находится маленький микронуклеус. Циста балантидия овальна, до 50—60 мкм в диаметре, покрыта двуслойной оболочкой, ресничек не имеет. Микронуклеус обычно не виден, а в цитоплазме отчетливо выделяется сократительная вакуоль (см. рис. 19.3, Д).
Балантидий может жить в кишечнике человека, питаясь бактериями и не принося ему вреда, но иногда внедряется в стенку кишки, вызывая образование язв с гнойным и кровянистым отделением. В этом случае в его цитоплазме часто обнаруживаются форменные элементы крови хозяина. Для заболевания характерны длительные поносы с кровью и гноем, а иногда и перфорация кишечной стенки с перитонитом. Как и при амебной дизентерии, В. coli может попадать в кровеносное русло и оседать в печени, легких и других органах, вызывая там образование абсцессов.
Особенностью этих инфузорий является их способность вырабатывать фермент гиалуронидазу, благодаря которой они внедряются и в неповрежденную стенку кишки, где на гистологических препаратах обнаруживаются целые скопления тканевых трофозоитов, морфологически не отличимых от живущих в просвете кишки, но не способных к образованию цист. Кроме человека, балантидий встречается также у крыс и свиней, которые и являются его основным резервуаром.
Лабораторная диагностика — обнаружение цист и трофозоитов в мазках фекалий больного.
Профилактика — как при лямблиозе, однако в связи с зоонозной природой балантидиаза следует также вести борьбу с грызунами и обеспечивать гигиеническое содержание свиней.
Кишечный балантидий.
Есть 2 ядра (вегетативное и генеративное), реснички. Деление осуществляется под оболочкой. Питается углеводной пищей. Если углеводов поступает достаточно – не переходит к тканевой форме. Язвы в кишечнике – 3-4 см. главный источник – домашние животные, продукты и вода, зараженные фекалиями крыс, свиней.
Инкубационный период длится до 15 лет. Обитают в сигмовидной, прямой, слепой кишках.
Диагностика – по обнаружению в фекалиях.
§
Тип Plathelmintes
Плоские черви имеют тело, уплощенное в дорсовентральном направлении. Полость тела отсутствует, внутренние органы погружены в рыхлую соединительную ткань — паренхиму. Кожно-мускульный мешок состоит из покровной ткани — тегумента, который представляет собой многоядерную неклеточную структуру, и трех слоев гладких мышц — продольных, поперечных и дорсовентральных. Движения, осуществляемые ими, медленны и несовершенны. Нервная система состоит из нервных узлов на переднем конце тела, от которых кзади отходят продольные нервные тяжи. Пищеварительная система, если она имеется, построена из глотки и кишечника, который слепо замкнут. Непереваренные остатки пищи выделяются через рот. Половая система гермафродитна и построена очень сложно.
Выделение осуществляется с помощью протонефридиальной системы, состоящей из отдельных выделительных клеток — протонефридиев. Они способны захватывать продукты диссимиляции и транспортировать их по внутриклеточным каналам, проходящим в их длинных отростках. Продукты экскреции поступают в собирательные трубочки, а оттуда либо непосредственно, либо через мочевой пузырь — во внешнюю среду.
Виды, имеющие медицинское значение, представлены в двух классах: Сосальщики и Ленточные черви.
Класс Сосальщики Trematoda
Класс Сосальщики включает около 4000 видов. Все они паразиты. Форма тела большинства сосальщиков листовидная. В процессе адаптации к паразитизму выработались мощные присоски — органы прикрепления. Многие сосальщики, кроме того, имеют мелкие шипики, покрывающие все тело и облегчающие им прикрепление к хозяину. Мелкие виды имеют пищеварительную систему в форме мешка или двух слепо замкнутых каналов. У крупных видов пищеварительная система сильно разветвлена и наряду с собственно пищеварением выполняет также транспортную функцию, перераспределяя по организму продукты пищеварения. В остальном сосальщики повторяют организацию плоских червей, описанных выше (рис. 20.1).

Рис. 20.1. Организация сосальщиков: 1—ротовая присоска, 2—мужские и женские половые отверстия; 3—семенники; 4—мочевой пузырь; 5—ветви кишечника, 6—желточники; 6—яичник; 8—брюшная присоска
Сосальщики резко обособлены от других плоских червей своеобразием жизненного цикла, в котором имеет место закономерное чередование поколений, способов размножения и хозяев (рис. 20.2). Половозрелая стадия всегда паразитирует в организме позвоночных животных. Выделяемое яйцо для успешного развития обычно должно попасть в воду. Из него выходит личинка — мирацидий, — снабженная светочувствительными глазками и ресничками, с помощью которых она свободно перемещается. Личинка обычно способна активно отыскивать промежуточных хозяев, используя фото-, гео- и хемотаксис.
Мирадиции попадают в организм брюхоногого моллюска определенного вида, строго специфичного для данного сосальщика. Здесь личинка превращается в материнскую спороцисту — стадию, претерпевшую в связи с паразитизмом наиболее глубокую дегенерацию. В ней развиты почти исключительно органы женской половой системы, благодаря которым она размножается партеногенетически. В результате этого образуются многоклеточные редии, которые также способны к партеногенезу. Таким образом, возможно формирование нескольких поколений редий. Последнее из них генерирует церкариев, покидающих организм моллюска и свободно плавающих в поисках основного или второго промежуточного хозяина.

Рис. 20.2. Цикл развития сосальщиков:
1—половозрелая форма в окончательном хозяине, 2—яйцо и мирацидий в воде, 3— личинки, размножающиеся партеногенетически в моллюске, 4—церкарий в воде, 5—метацеркарий во втором промежуточном хозяине
В первом случае церкарий либо самостоятельно внедряются в кожу хозяина, либо, инцистируясь на растениях, оказываются проглоченными травоядными животными или человеком. Во втором случае церкарий отыскивают животных, использующихся основными хозяевами для питания, и образуют в них покоящиеся стадии — инцистированные метацеркарий. Церкарий в отличие от мирацидиев не обладают хемотаксисом, а использование только гео- и фототаксиса, а также инцистирование на траве не позволяют им находить специфичных хозяев. Поэтому основная масса церкариев погибает либо не найдя хозяев вообще, либо попав в организмы таких видов, развитие в которых невозможно. После проникновения инвазионных стадий сосальщиков в основного хозяина они мигрируют у него в организме и находят тот орган, где достигнут половой зрелости и будут обитать всю последующую жизнь.
Отсутствие свободноживущих видов в классе Сосальщики на фоне общего примитивизма их организации, сложный цикл развития, в котором участвуют специфические промежуточные и основные хозяева, огромная плодовитость, достигающаяся разными способами размножения, и сохранившаяся связь с водной средой обитания свидетельствуют о древности паразитического образа жизни этих организмов. Среди хозяев сосальщиков наиболее специфичными являются именно моллюски: для каждого вида паразита, обитающего на стадии мариты нередко у нескольких видов основных хозяев, первым промежуточным хозяином практически всегда является только один вид улиток.
Это означает, что на протяжении эволюции между сосальщиками и моллюсками возникли наиболее совершенные взаимные адаптации. Моллюски были, вероятно, первыми и единственными хозяевами сосальщиков еще в период, предшествующий возникновению позвоночных. Об этом свидетельствует также сохранившийся в цикле развития сосальщиков партеногенез — рудиментарная форма полового размножения, — проходящий именно в организме моллюсков. Наряду с этими чертами в цикле развития имеются такие признаки, которые указывают на рекапитуляции свободного образа жизни: обязательный выход яиц во внешнюю среду и наличие активно плавающих расселительных стадий с органами чувств — мирацидиев и церкарий.
Возникновение и дальнейший биологический и морфофизиологи-ческий прогресс позвоночных, освоивших все благоприятные для жизни среды, открыли и сосальщикам широкие возможности адаптивной эволюции за счет приспособления к новым хозяевам. Адаптации сосальщиков к позвоночным как к новым средам обитания не ограничились только использованием их широкого видового разнообразия. Сосальщики адаптировались к обитанию в самых разнообразных органах, тканях и системах — от кожных покровов и органов чувств до кровеносных сосудов внутренних органов.
Однако большинство сосальщиков-паразитов человека обитает в пищеварительной системе; некоторые виды живут в легких, другие — в кровеносных сосудах брюшной полости и малого таза. Человек заражается сосальщиками в зависимости от вида разными способами: при контакте с водой и проникновении церкарий через кожу, при поедании продуктов животного происхождения с метацеркариями и при употреблении растений в пищу, если на их листьях инцистированы церкарии паразитов.
После попадания в организм человека большинство сосальщиков осуществляют сложные миграции по пути к органам своей окончательной локализации. Миграция происходит по кровеносным сосудам, непосредственно по пространствам между органами и по полости тела. Во время миграции сосальщики вызывают у хозяина тяжелые интоксикации и аллергические состояния, но диагностировать заболевание в этот момент крайне сложно. Заболевания, вызываемые сосальщиками, называют трематодозами.
Для диагностики трематодозов используют методы обнаружения яиц в фекалиях, моче или мокроте в зависимости от локализации паразитов, а также аллергические пробы.
В связи с тем что сосальщики, обитающие у человека, поражают также и ряд других видов млекопитающих, соответствующие трематодозы относят к природно-очаговым зоонозным заболеваниям, поэтому их полная ликвидация практически невозможна.
В зависимости от особенностей цикла развития сосальщиков, паразитирующих у человека, можно разделить на следующие группы: 1) развивающиеся с одним промежуточным хозяином и обитающие в пищеварительной системе; 2) развивающиеся с одним промежуточным хозяином и обитающие в кровеносных сосудах; 3) развивающиеся с двумя промежуточными хозяевами.
§
Характеристика:
трехслойность;
кожно-мускульный мешок;
отсутствие полости тела;
билатеральная симметрия;
тело сплюснуто в дорсовентральном направлении;
наличие развитых систем органов (половая, нервная, мышечная, пищеварительная, выделительная);
отсутствуют кровеносная и дыхательная системы;
наличие органов прикрепления;
первичноротые.
Класс СОСАЛЬЩИКИ TREMATODA(более 4000 видов).
Все они ведут паразитический образ жизни. Во взрослой стадии поселяются в теле позвоночных, в т.ч. человека, в личиночных стадиях поселяются в беспозвоночных, чаще всего в моллюсках. Большинство обитают в пищеварительной системе и железах, связанных с ЖКТ. Некоторые живут в легких, мочеполовых органах, реже поражают головной мозг. Ни одна другая паразитическая группа не распространилась по телу человека так широко, как плоские черви и не приспособилась так хорошо к условиям жизни в организме хозяина. Приспособления связаны с особенностями строения и цикла развития. Важное приспособление – огромная плодовитость, способность размножаться партеногенетическим путем. Среди сосальщиков много опасных паразитов человека и животных. Поселяясь в организме хозяев, они вызывают серьезные заболевания, паразитирование очень плохо сказывается на здоровье хозяина, возможен летальный исход. Заболевания носят название трематозов.
Тело обычно плоское, листовидное, не расчленяется. Имеются 2 присоски – ротовая и брюшная (поэтому иногда сосальщиков называют по старинке двуустками). Окраска тела невзрачная, грязно-белая, сероватая, розоватая, иногда с темными пятнами. Половозрелая гермафродитная особь называется – марита. Стенку ее составляет кожно-муск5ульный мешок, состоящий из тегумента и мышечного слоя. Тегумент – клетки, объединившие цитоплазму с ядрами внутри. Снаружи расположены кольцевые мышцы, затем идут слои продольных и диагональных мышц.
Пищеварительная система состоит из глотки, пищевода и двух ветвей кишечника, которые могут быть трубчатыми разветвленными или неразветвленными. Заканчивается кишечник слепо, остатки пищи удаляются через ротовое отверстие.
Выделительная система имеет центральный выделительный канал, от которого отходят канальцы, конечная часть – терминальные клетки. Есть специальные реснички, которые собирают жидкость и двигают ее в направлении канальцев, а затем в главный выделительный канал, который открывается выделительной порой.
Нервная система представлена окологлоточным нервным кольцом и 3 парами нервных стволов, отходящими в разные стороны тела.
Половая система. Занимает большую часть тела. Мужской половой аппарат просто устроен. Есть 2 семенника, которые могут иметь различную форму: лопастные, розетковидные и др. различается также и их положение в теле особи. Также есть 2 семяпровода, которые идут в копулятивный орган, в него же открывается семяизвергательный канал. Женская половая система устроена сложнее. Есть яичник (разной формы), яйцевод, который ведет в оотип, где происходит формирование яиц, далее располагается матка. Оотип имеет протоки 2х желточников и проток железы Мелиса. Оплодотворение яйцеклеток происходит в оотипе, там же происходит формирование яиц. Постепенно созревая, яйца перемещаются в матку и выводятся наружу через половое отверстие. Матка открытого типа.
Печеночный сосальщик. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
Класс Trematoda
Печеночный сосальщик Fasciola hepatica (рис. 20.3) — возбудитель фасциолеза. Тело паразита листовидное, передний конец клювообразно оттянут. Матка невелика и розеткой располагается позади брюшной присоски. Кзади от матки находятся сильно разветвленные семенники, яичники, желточники и ветви кишечника. Яйца крупные, желтовато-коричневого цвета. Фасциолез встречается чаще в странах с теплым влажным климатом.
Жизненный цикл печеночного сосальщика типичен для этой группы паразитов. Окончательные хозяева паразита — крупные травоядные млекопитающие и человек. Промежуточный хозяин — малый прудовик Lymnea truncatula. Интересно, что на территории Австралии нет моллюсков этого вида, а фасциолез распространен широко. Этот паразит, будучи занесенным сюда вместе со скотом, адаптировался к другому промежуточному хозяину — L. tomentosa. Заражение основного хозяина происходит при поедании травы с заливных лугов. Случаи заражения человека связаны с употреблением щавеля и особенно часто — водяного кресса — полуводного растения, широко употребляющегося в пищу в Западной Европе. В кишечнике основного хозяина личинки освобождаются от оболочек, пробуравливают его стенку и попадают в полость брюшины, откуда мигрируют в печень, проникают через печеночную ткань в жёлчные ходы, где через 3—4 мес. достигают половой зрелости и начинают откладывать яйца.
Печеночный сосальщик гигантский Fasciola gigantica отличается от предыдущего вида большими размерами (до 75 мм) и слабым выступанием вперед передней части тела. Цикл развития, диагностика и профилактика не отличаются от предыдущего вида. В связи с крупными размерами этого вида фасциолез, вызываемый им, протекает более тяжело. У человека фасциолез, вызываемый гигантским печеночным сосальщиком, встречается в Юго-Восточной Азии, на Гавайских о-вах, в Узбекистане.
· Гермафродит
· повсеместно
· пром хозяин — малый прудовик
· инвазионная стадия промеж — мирацидий
· личиночные стадии — спороциста, редии, циркарии
· окончательный хозяин — КРС, человек
· инвазионная стадия окончат — адолексария
· пути заражения: проглатывание адолексарий с водой, с овощами, помыти водой из загрязненного водоёма
· обнаружение яиц в фекалиях
· не пить воду их рек и прудов, не мыть овощи речной водой, уничтожение моллюсков
· локализация: желчные протоки печени
Фасциолопсис. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики
Тип Plathelmintes
Класс Trematoda
Fasciolopsis busky – возбудитель фаслопсидоза. Толстое тело, неразветвленные кишечные трубки. Взрослые формы обитают в тонком кишечнике. Промежуточные хозяева – водные моллюски. Церкарии инцистируются на плодах водного ореха и водяного каштана. Вызывает хронические желудочно-кишечные расстройства, сильно истощает больных.
Кошачий сосальщик. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики. Очаги описторхоза.
Тип Plathelmintes
Класс Trematoda
Кошачий сосальщик Opisthorchis felineus (рис. 20.5, В) — возбудитель описторхоза. Длина тела до 13 мм. Характерная особенность—два хорошо окрашивающихся лопастевидных семенника на заднем конце тела. Яйца длиной 26—30 мкм, с крышечкой. Описторхоз — эндемичное для России заболевание. Встречается у человека наиболее часто в Западной Сибири, но изредка проявляется и в европейской части СНГ — в Волжско-Камском бассейне, в бассейне рек Дона, Днепра, Днестра и Северского Донца. Обнаружен и в бассейне Немана. Известны природные очаги без участия человека также в Казахстане.
Первый промежуточный хозяин кошачьего сосальщика — моллюск Bithynia leachi, второй — карповые рыбы, в мышцах которых локализуются метацеркарии паразита. Окончательные хозяева — различные дикие и домашние рыбоядные млекопитающие и человек.
Особенностью паразитов этой группы является миграция личиной в организме человека, поэтому на первом этапе течения заболевания проявляются аллергические реакции. Длительное паразитирование большого количества этих сосальщиков в желчных ходах нарушает отток желчи, кровоснабжение и дает осложнения в 15—20% случаев цирротическими изменениями печени, а иногда и злокачественным перерождением печеночной ткани — возникновением первичного рака печени.
При диагностике заболеваний следует исследовать фекалии для обнаружения яиц, а также провести дуоденальное зондирование, при котором возможно выделение через зонд не только яиц, но и половозрелых паразитов.
Эуритрема (сосальщик поджелудочной железы). Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики
Тип Plathelmintes
Класс Trematoda
Сосальщик поджелудочной железы Eurytrema pancreaticum (рис. 20.5, Ж) — возбудитель эуритрематоза. Тело сильно расширено, длиной до 15мм. Характерны очень крупные присоски и матка, лежащая на заднем конце тела. Яйца до 50 мкм длиной. Паразит у животных встречается в Южной и Юго-Восточной Азии и в южных районах СНГ — в Казахстане, Киргизии, на Дальнем Востоке. .
Первый промежуточный хозяин паразита — моллюск из р. Sradybaena, второй — луговой кузнечик р. Conocephalus или сверчок р. Oecanthus. Человек заражается, употребляя в пищу кузнечиков, не прошедших термической обработки, поэтому заражение человека встречается только в странах Юго-Восточной Азии. Сосальщик поселяется в протоках поджелудочной железы и вызывает симптомы хронического панкреатита.
Диагностика — обнаружение яиц в фекалиях и дуоденальном содержимом при зондировании.
Профилактика заражения человека — отказ от поедания насекомых или их предварительная термическая обработка.
Ланцетовидный сосальщик. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
Класс Trematoda
Эта группа сосальщиков интересна своеобразием адаптации к среде обитания. Яйца паразитов должны попасть на почву или растения. Они содержат зрелых мирацидиев, которые из яиц не выходят до тех пор, пока они не будут проглочены наземными моллюсками — их первыми промежуточными хозяевами. Типичные подвижные церкарии этими сосальщиками не образуются. Они выделяются моллюсками в виде слизистых комочков на растения, где и поедаются вторыми промежуточными хозяевами — насекомыми. Насекомые, пораженные метацеркариями этих паразитов, становятся малоподвижными и могут оказаться съеденными окончательными хозяевами — травоядными животными и человеком. Как и большинство сосальщиков, эти паразиты в организме окончательного хозяина осуществляют миграцию.
Особенности цикла развития позволяют этим сосальщикам расселиться очень широко и заселить безводные зоны с сухим климатом. В связи с тем что насекомые весьма редко попадают в пищу к человеку, заболевания человека нечасты. Однако в Юго-Восточной Азии культурные традиции допускают питание насекомыми. Поэтому на территории Лаоса, Таиланда, Южного Китая и Японии заболевания, вызванные этими паразитами, встречаются чаше, чем в других зонах.
Ланцетовидный сосальщик Dicrocoelium lanceatum (рис. 20.5, Е) — возбудитель дикролоцелиоза — паразит, внешне напоминающий кошачьего сосальщика размерами и формой, но семенники у него имеют более округлую форму и расположены на передней стороне тела. Яйца его несколько крупнее — до 45мкм. Распространен повсеместно.
Первый промежуточный хозяин — моллюски р. Helicella или Zebrina, второй — муравей р. Formica. Человек заражается случайно, проглатывая инвазированного муравья. Поселяется этот сосальщик в жёлчных ходах печени.
Диагностика — как при всех трематодозах с поражением печени.
Личная профилактика — необходимо следить, чтобы в пищу не попадали муравьи. Общественная профилактика — дегельминтизация скота и санитарная охрана пастбищ.
Ланцетовидный сосальщик.
Окончательный хозяин – травоядные животные. Яйца и мирацидии выделяются во внешнюю среду, образуются сборные цисты, которые съедаются моллюсками. Происходит поражение центрального нервного ганглия, следовательно, нарушение движений и гибель.
Легочный сосальщик. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
Класс Trematoda
р. Paragonimus
Легочный сосальщик Paragonimus westermani (рис. 20.5, Д) — наиболее часто встречающийся возбудитель парагонимоза. Кроме него известно еще пять видов сосальщиков, чаще паразитирующих у животных, но поражающих также и человека. Основной вид распространен по всему тропическому поясу Старого и Нового Света, исключая Австралию, а за пределами тропиков — в Дальневосточном регионе, включая южные районы Приморского края и Приамурья. Остальные виды описаны на ограниченных ареалах — в Японии, Южном Китае, Западной Африке.
Легочные сосальщики имеют необычную для сосальщиков форму тела: они напоминают семя апельсина и имеют размеры до 12 мм. Яйца до 0,118 мм длиной.
Первый промежуточный хозяин — моллюски из родов Semisulcospira, Oncomelania и некоторых других. Второй промежуточный хозяин — крабы из р. Eriocheir, Potamon, раки родов Cambarus, Procambarus, а также креветки р. Macrobrachium. Окончательные хозяева — человек и животные, питающиеся ракообразными, — выдры, норки, свиньи, кошки, собаки и некоторые грызуны.
Для диагностики заболевания необходимо исследовать мокроту больных, в которой обнаруживаются яйца, а также фекалии, куда яйца могут попадать при проглатывании мокроты. В пищеварительном тракте они не изменяются.
Личная профилактика парагонимоза заключается в отказе от поедания сырых ракообразных. Общественная профилактика соответствует мерам, применяемым против сосальщиков, развивающихся в водной среде с двумя промежуточными хозяевами.
Возбудитель парагонимоза. Паразитирует в легких свиней, человека и других млекопитающих. Тело красноватое, коричневатое. 5-7 мм в ширину, 12 в длину. Размеры яиц 120х50. Яйца с крышечкой, дозревают в воде, выходят мирацидии, поселяющиеся в моллюске (стадии спороцисты, редии, церкарии). Церкарии попадают в пресноватых раков и крабов, внедряясь в тело. Из церкариев формируются метацеркарии – инвазионная стадия для основного хозяина. Попав в организм основного хозяина, метацеркарий прободает стенку кишечника, попадает в брюшную полость, прободает диафрагму и достигает легких. В легких вокруг паразита формируется полость, заполненная продуктами распада тканей и метаболизма паразита. Парагонимоз распространен в Китае, Корее, Африке, Дальнем Востоке, Японии. Иногда паразиты попадают в мозг, вызывая энцефалит и менингоэнцефалит. На начальной стадии определяются только иммунологически, в хронической стадии паразитов можно обнаружить паразитологически.
146.Китайский сосальщик. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
Класс Trematoda
§
Тип Plathelmintes
Класс Trematoda
Возбудители шистосоматоза. Живут в кровеносных сосудах, причем чаще в венах
Урогенетальная шистосома. Самка 10-15 мм, толщина 1 мм, самец в 2 раза длинней и тоньше. У самцов есть гинекоморфный канал, где находятся самки при спаривании. Мирацидий выходит из яйца при температуре воды 15-20 градусов и рН больше 7. необходима также хорошая аэрация. Затем попадают в моллюска, из них выходят церкарии, которые активно внедряются в тело окончательного хозяина (человек, высшие обезьяны). Личинки мигрируют по лимфатическим путям в легкие, печень, преимущественно оказываясь в венозных сплетениях мочевого пузыря и мочеполовых путях. Яйца без крышечки, имеют шип на конце. Мирацидий в яйце выделяется протеолитические ферменты. С помощью шипа он прободает кровеносный сосуд и попадает в мочевой пузырь.
Наиболее часто заражаются сельскохозяйственные рабочие, рыбаки, лодочники – до 50-60% . Имеют место токсические и аллергические реакции. Тяжелое заболевание. Внедрение церкариев, паразитирующих в птицах, вызывает зуд и крапивницу – шистосомный дерматит.
К этой группе относятся так называемые кровяные сосальщики — шистосомы. Это раздельнополые организмы. Самцы имеют широкое тело, а самки — шнуровидное и в половозрелом состоянии находятся в гинекофорном канале на брюшной стороне самцов. Присоски невелики и располагаются на переднем конце тела. Все кровяные сосальщики обитают в тропических широтах Азии, Африки и Америки.
В связи с тем что половозрелые шистосомы живут в кровеносных сосудах, яйца, которые откладывают самки, имеют специальные приспособления для выведения в полостные органы и далее во внешнюю среду: они снабжены шипами, через которые выделяются ферменты, растворяющие ткани хозяина. За счет этих ферментов яйца пробуравливают стенки сосудов, проникают в ткани, могут попадать в кишечник или мочевой пузырь в зависимости от вида паразита. Опасным и характерным для этих паразитов является гематогенный занос яиц в различные органы, где вокруг них возникают локальные воспалительные процессы.
Как и у всех сосальщиков, личиночные стадии, размножающиеся партеногенезом, развиваются в водных моллюсках. Характерными являются церкарии — они имеют раздвоенный хвост, а на переднем конце — железы проникновения, с помощью которых проникают через кожу в кровеносную систему окончательного хозяина при нахождении его в воде (см. рис. 18.7). При этом они вызывают кожные поражения — церкариозы, выражающиеся в появлении сыпи, зуде и других аллергических проявлениях. При массовом попадании церкарии в легкие возникает пневмония.
Церкариозы могут вызываться не только шистосомами, паразитирующими у человека, но и несколькими видами кровяных сосальщиков, обитающих у водоплавающих птиц. Эти виды церкарии у человека обычно погибают либо в коже, либо в легких.
Личинки шистосом, патогенных для человека, мигрируют по организму и оседают в венах брюшной полости и малого таза, где и достигают половой зрелости. Патогенное действие половозрелых шистосом выражается в токсико-аллергических реакциях хозяина и местных проявлениях: характерны кровотечения из пораженных органов, образование изъязвлений и полипов, склонных к злокачественному перерождению.
Диагностика заключается в обнаружении яиц шистосом в фекалиях или моче. Проводят также аллергические внутрикожные пробы и иммунобиологические реакции в пробирке.
Профилактика шистосоматозов: необходимо остерегаться длительных контактов с водой в зонах распространения этих паразитов. В связи с тем что церкарии могут проникать и через слизистые оболочки, для питья следует использовать только обеззараженную воду. В целях общественной профилактики необходима охрана водоемов от загрязнения необеззараженными сточными водами. В ряде случаев возможна также борьба с моллюсками — промежуточными хозяевами паразитов — разными способами. В Бразилии эффективным оказался метод биологической борьбы с шистосомами — использование рыбки гуппи — Lebistes reticulatus, которая активно поедает церкарий, выходящих из зараженных моллюсков.
Для профилактики церкариозов необходимо воздержаться от купания в пресноводных водоемах, в которых обитают водоплавающие птицы.
У человека часто паразитируют три вида шистосом, распространенных в тропиках Азии, Африки и Америки. В Центральной и Южной Африке описаны случаи инвазии человека еще четырьмя видами, обычно паразитирующими у животных. Кожные поражения в виде церкариозов могут наблюдаться в любой климатической зоне, где обитают водоплавающие птицы — окончательные хозяева ряда видов шистосом.
Schistosoma haematobium (рис. 20.4, А) — возбудитель мочеполового шистосоматоза. Самец длиной до 1,5, а самка — до 2,0 см. Поверхность тела мелкобугристая. Яйца очень крупные, размером до 0,16 мм; на конце находится длинный шип.
Промежуточными хозяевами являются моллюски из р. Bullinus, Planorbis или Planorbarius; окончательными хозяевами — человек и обезьяны, у которых паразит после миграции поселяется в венах мочевого пузыря и органах половой системы.
Встречается от Африки до Юго-Западной Индии.
Характерным для мочеполового шистосоматоза является гематурия (кровь в моче), боли в надлобковой области, нередко образование камней в мочевыводящих путях. В зонах распространения этого заболевания рак мочевого пузыря встречается в 10 раз чаще, чем в областях, свободных от шистосоматоза.
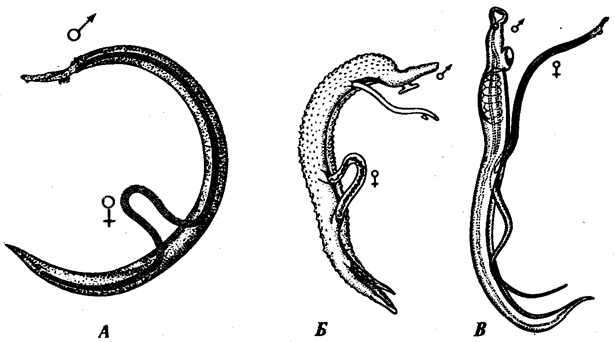
Рис. 20.4. Кровяные сосальщики.
А — Schistosoma haematobium; Б — Sch..mansoni; В — Sch. japonicum
При диагностике обнаруживают яйца в моче, а также характерные изменения мочевого пузыря и влагалища: изъязвления, полипозные разрастания и местные воспалительные процессы.
Schistosoma mansoni (рис. 20.4, Б) — возбудитель кишечного шистосоматоза. В отличие от предыдущего вида имеет несколько меньшую длину (до 1,6 мм) и крупнобугристую поверхность тела. Яйца по размерам соответствуют яйцам Sch. haematobium, но шип находится на боковой поверхности. Промежуточными хозяевами являются моллюски р. Biomphalaria, а окончательными — человек, обезьяны, собаки и грызуны. Ареал распространения шире, чем у предыдущего вида: он охватывает Северную, Экваториальную и Юго-Восточную Африку, Юго-Западную Азию; паразит занесен и в Западное полушарие — в Бразилию, Венесуэлу, Гайану и на Антильские о-ва.
У человека заселяет брыжеечные вены толстого кишечника и систему воротной вены печени. В связи с этим поражения возникают в первую очередь в толстом кишечнике (явления колита, понос с примесью крови, возможен полипоз толстой кишки) и в печени (венозный застой и цирроз).
При диагностике обнаруживают яйца в фекалиях.
Schistosoma japonicum (рис. 20.4, В) — возбудитель японского шистосоматоза. Этот паразит по размерам не отличается от Sch. haematobium, но поверхность его тела совершенно гладкая. Яйца более округлые, чем у описанных видов, а шип, расположенный на боковой поверхности, имеет очень малые размеры. Промежуточные хозяева — моллюски р. Oncomelania, а окончательные — человек и большое количество видов диких и домашних млекопитающих — грызуны, собаки, копытные. Ареал охватывает Восточную и Юго-Восточную Азию.
Паразит локализуется в венах кишечника, поэтому проявления болезни и диагностика соответствуют описанному кишечному шистосоматозу. Распространение яиц паразита по кровеносным сосудам в различные органы, в том числе в головной мозг, встречается чаще, чем при других формах шистосоматоза.
· раздельнополые
· средиземноморье, ближний восток, африка
· пром хозяин — моллюски рода planorbis и bullineus
· инвазионная стадия промеж — мирацидий
· личиночные стадии — спороциста 1 порядка, 2 порядка, циркарии
· окончательный хозяин —обеьяны, человек
· инвазионная стадия окончат — церкария
· активное внедрение церкарий через кожу, проглатывание с водой
· обнаружение яиц в моче
· не купаться в зараженных водоёмах, не пить воду из них
- вены и венулы малого таза
148.Бычий цепень. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Класс Ленточные черви Cestoidea
Как и сосальщики, все ленточные черви — паразиты, главным образом позвоночных животных. Класс насчитывает около 3500 видов. Форма тела этих червей лентовидная. У большинства видов тело, или стробила, разделено на многочисленные членики — проглоттиды. На переднем конце находится головка, или сколекс, несущая органы прикрепления — присоски, крючья или присасывательные щели — ботрии. За головкой следует несегментированная шейка, от которой сзади постепенно отпочковываются молодые проглоттиды. В них системы органов не дифференцированы. В средней части стробилы лежат членики с развитой мужской и женской половыми системами. Они называются гермафродитными. Последние проглоттиды стробилы содержат почти исключительно матку, заполненную яйцами, и рудименты остальных органов. Эти членики называются зрелыми. В процессе роста червя задние, зрелые, членики постепенно отрываются, а от шейки образуются все новые, молодые проглоттиды.
Пищеварительная система у ленточных червей отсутствует в связи с длительной эволюцией в условиях паразитизма. Питание осуществляется всей поверхностью тела за счет пиноцитоза тегументом. Нервная система и органы выделения построены по плану, характерному для всего типа плоских червей. Половая система состоит из тех же органов, что и у сосальщиков, но представлена в каждом ленточном черве в огромном количестве копий, соответствующих количеству проглоттид (рис. 20.6).
Цикл развития ленточных червей отражает их более глубокие адаптации к паразитизму по сравнению с сосальщиками. Об этом свидетельствует то, что свободноживущие расселительные стадии имеются только в одной группе этих червей — лентецов, считающихся наиболее древними по происхождению. Большинство ленточных червей попадают во внешнюю среду только в виде яйца, но размножаются лишь в организме хозяина. Некоторые наиболее специализированные паразиты способны обеспечивать аутоинвазию хозяина с помощью яиц, даже не выходящих во внешнюю среду.
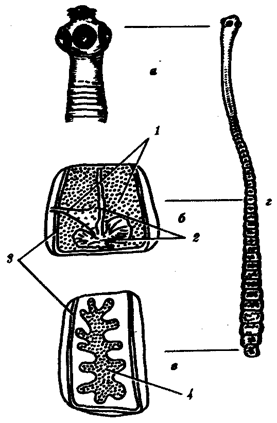
Рис. 20.6. Организация ленточных червей:
а—головка; б—гермафродитный членик; в—зрелый членик; г—стробила;
1—мужские половые органы; 2—женские половые органы; 3—каналы выделительной системы; 4—матка со зрелыми яйцами
Цикл развития наиболее древних ленточных червей связан с водной средой, самые примитивные ленточные черви и близкие им формы являются эктопаразитами наиболее древних позвоночных — хрящевых рыб, обитая у них на кожных покровах, жабрах, а также в глотке и переходя, таким червей: образом, к эндопаразитизму. Появление и быстрый расцвет более прогрессивных позвоночных – костных рыб, земноводных и т.д. — привел и ленточных червей к освоению новых сред обитания вместе с новыми хозяевами, в том числе с наземными позвоночньми. Современные ленточные черви в цикле развития имеют две стадии: половозрелую и личиночную. Половозрелая стадия паразитирует обычно в тонком кишечнике позвоночных. Личиночная стадия, или финна, — тканевый паразит в организме промежуточных хозяев, в основном позвоночных, но иногда также членистоногих (рис. 20.7).
С фекалиями окончательного хозяина яйца паразитов попадают во внешнюю среду. Они содержат личинку — онкосферу, которая будет развиваться уже в промежуточном хозяине при попадании яиц в его пищеварительную систему. Здесь онкосфера с помощью крючьев проникает через кишечную систему в кровеносное русло или лимфатические сосуды и мигрирует по организму, оседая в печени, легких, мышцах, центральной нервной системе и т. д. Разрастаясь, она превращается в финну.
Финна — промежуточная форма паразита, обычно с полостью внутри и со сформировавшейся головкой. Финны некоторых ленточных червей способны размножаться бесполым путем за счет внутреннего или наружного почкования. Это увеличивает вероятность заражения основного хозяина и выживания потомства паразита. Окончательный хозяин, обычно хищное животное или человек, проглатывает финну вместе с тканями промежуточного хозяина. При этом головка паразита прикрепляется к стенке кишки хозяина и начинается рост шейки, образование члеников и развитие гельминта.
Рис. 20.7. Цикл развития ленточных червей:
1 — половозрелая стадия в кишечнике окончательного хозяина, 2 — яйцо по внешней среде, 3 — финнозная стадия в тканях промежуточного хозяина
В связи с тем что окончательный хозяин заражается этими гельминтами, поедая промежуточных хозяев, взаимоотношения хозяина и паразита на разных стадиях развития червей различны.
Если основной хозяин, в кишечнике которого находится гельминт, страдает от него относительно несильно, то жизнеспособность промежуточного хозяина с финнами этих червей в легких, мозгу или печени оказывается резко снижена. Это увеличивает вероятность именно инвазированных организмов быть съеденными окончательным хозяином.
По особенностям биологии ленточных червей, имеющих медицинское значение, можно разделить на группы, жизненный цикл которых связан и не связан с водной средой. Вторая группа подразделяется на гельминтов: а) использующих человека как окончательного хозяина, б) обитающих в человеке как в промежуточном хозяине, в) проходящих в человеке весь жизненный цикл.
Соответственно этому пути заражения человека патогенное действие паразитов, диагностика, лечение и профилактика соответствующих заболеваний различны.
Болезни, вызываемые ленточными червями, называют цестодозами. Многие виды ленточных червей поражают только человека, другие встречаются также и в природной обстановке, для них характерно существование классических природных очагов. Цестод, обитающих в человеке как основном хозяине, сближает то, что они живут в кишечнике и всегда в небольшом количестве. Это объясняется выраженной внутривидовой конкуренцией, в которой выживают лишь единичные особи. На интенсивности размножения это не отражается, так как плодовитость их огромна.
Бычий цепень Taeniarrhynchus saginatus (рис. 20.8, Б) — возбудитель тениаринхоза, достигает в длину 4—10 м. На головке имеет только четыре присоски. Гермафродитные членики квадратной формы, матка в них не разветвляется, а яичник состоит из двух долей. Зрелые членики сильно вытянуты. Матка очень разветвлена, число ее боковых ветвей достигает 17—34 пар. Яйца содержат онкосферы, расположенные под тонкой прозрачной оболочкой, которая быстро разрушается. Онкосферы имеют три пары крючьев и толстую, радиально исчерченную оболочку. Диаметр онкосфер около 10 мкм. Тениаринхоз распространен повсеместно, где население употребляет в пищу сырое или недостаточно обработанное говяжье мясо.
Жизненный цикл бычьего цепня типичен. Основной хозяин только человек, промежуточный — крупный рогатый скот. Особенностью этого вида является то, что с фекалиями человека членики выделяются группами по 5—6. Корова, проглотив такие членики, становится промежуточным хозяином паразита. В ее мышцах формируются финны, называемые цистицерками. Финна представляет собой пузырек, заполненный жидкостью, в котором находится сколекс. В мышцах финны могут сохранять жизнеспособность долгие годы. При поедании мяса такой коровы в желудке под действием кислой среды желудочного сока головка вывертывается, прикрепляется к стенке кишки и развивается новый цепень.
Диагностика проводится несложно — при обнаружении зрелых члеников в фекалиях, так как членики имеют характерное строение.
Профилактика тениаринхоза заключается в охране пастбищ от заражения фекалиями человека.
Повсеместно
КРС (яйцо с онкосферой) — онкосфера, финна типа цистицерк
Человек (финна типа цистицерк)
при поедании финнозной говядины, не прошедшей термическую обработку
тонкий кишечник
термическая обработка мяса
Свиной цепень. Систематическое положение, морфология, цикл развития, географическое распространение, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
§
Тип Plathelmintes
класс Cestoidea
Свиной цепень Taenia solium (рис. 20.8, В) — возбудитель тениоза и цистицеркоза. Этот паразит меньше предыдущего, он достигает в длину 3 м. На головке кроме присосок у него находится венчик из 22—32 крючьев. В гермафродитных члениках не две, а три дольки яичника; матка в зрелых члениках имеет не более 12 пар боковых ответвлений. Яйца не отличаются от яиц предыдущего вида.
Жизненный цикл свиного цепня типичен. Окончательный хозяин паразита — человек. Характерной особенностью является способность члеников активно выползать из заднепроходного отверстия поодиночке. При подсыхании оболочка их лопается и яйца могут свободно рассеиваться во внешней среде. Этому процессу могут способствовать птицы и мухи. Из яиц, проглоченных промежуточным хозяином — свиньей, развивается онкосфера и позже цистицерки, как и у предыдущего вида.
Промежуточными хозяевами этого гельминта кроме домашних и диких свиней могут быть кошки, собаки и человек: В этом случае у них, так же как и у свиней, развивается цистицеркоз. Человек может проглотить яйца свиного цепня случайно, но более часто цистицеркоз возникает как осложнение тениоза. При этом заболевании особенно часто возникает обратная перистальтика кишечника и рвота. Зрелые членики могут таким образом попасть в желудок, перевариться там, а освободившиеся онкосферы проникают в сосуды кишечника, разносятся кровью и лимфой по организму, где в печени, мышцах, легких, мозге и других органах формируются цистицерки. Это может привести к быстрому смертельному исходу.
Лабораторная диагностика тениоза основана на обнаружении характерных зрелых члеников в фекалиях; диагностика цистицеркоза сложнее — путем рентгенологического обследования и постановки иммунологических реакций.
Для личной профилактики тениоза необходимо термически обрабатывать свинину, а цистицеркоза — соблюдать правила личной гигиены. Общественная профилактика — закрытое содержание свиней.
· Повсеместно
· КРС (яйцо с онкосферой) — онкосфера, финна типа цистицерк
· Человек (финна типа цистицерк)
· при поедании финнозной свинины, проглатывание зрелых яиц или заброс зрелых члеников при антиперильстальтических движениях кишечника
· тениоз: тонкий кишечник, цистицеркоз: глаза, мышцы, гм
- термическая обработка мяса
Карликовый цепень. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
класс Cestoidea
Циклы развития, проходящие полностью в одном хозяине, встречаются не часто среди многоклеточных паразитов. Способность гельминта размножаться в хозяине длительное время, не покидая его на стадии яйца, — явление крайне редкое. Таким необычным паразитом человека стал карликовый цепень Hymenolepis nana — возбудитель гименолепидоза (рис. 20.11). Это мелкий гельминт длиной до 5 см с головкой, снабженной крючьями и присосками. Зрелые членики столь нежны, что, развиваясь, полностью разрушаются еще в кишечнике, поэтому яйца быстро выходят из них и попадают в фекалии. Размер яиц до 40 мкм. Они имеют округлую форму и бесцветны.

Рис. 20.11. Карликовый цепень
Гименолепидоз встречается повсеместно, особенно в странах с сухим и жарким климатом. Поражаются преимущественно дети.
Жизненный цикл этого паразита за время длительной адаптации к организму человека претерпел существенные изменения. Если его яйца, выделившись из организма с фекалиями, попадают в пищеварительную систему мучного хруща из р. Tenebrio, в нем разовьется финнозная стадия червя — цистицеркоид. При проглатывании инвазированного жука с непропеченным тестом в кишечнике человека из цистицеркоидов разовьются взрослые паразиты. В современных условиях такое явление происходит крайне редко. Если человек проглотит яйца карликового цепня при несоблюдении правил личной гигиены, то в ворсинках тонкого кишечника из них развиваются цистицеркоиды, которые позже, разрушая ворсинки, выпадают в просвет кишечника и превращаются в половозрелых гельминтов. Яйца этого паразита могут не выделяться во внешнюю среду вовсе, быстро достигая зрелости еще в кишечнике. В таком случае из них развиваются цистицеркоиды, как и в предыдущем случае, а затем и взрослые цепни.
Таким образом, у одного и того же вида сосуществуют три варианта развития: типичный для ленточных червей со сменой двух хозяев; с утратой промежуточного хозяина; вообще без выхода во внешнюю среду даже на стадии яйца. Эти три варианта соответствуют трем этапам эволюции биологии карликового цепня в процессе адаптации его к паразитированию в таком специфическом хозяине, как человек. Способность этого паразита к аутоинвазии хозяина даже без выхода во внешнюю среду обусловливает длительность заболевания на фоне смены поколений паразитов и сложность полного излечения в связи с тем, что в организме человека постоянно сосуществуют как личиночные формы в ворсинках, так и половозрелые — в просвете кишки. Однако в связи с тесным контактом цистицеркоидов карликового цепня с тканями хозяина у человека возникает иммунитет против них. Он затрудняет развитие следующих поколений паразитов, обеспечивая отторжение и гибель все большего количества цистицеркоидов, формирующихся в процессе аутоинвазии.
Таким образом, несмотря на аутоинвазию, гименолепидоз не может продолжаться бесконечно. После смены нескольких поколений паразитов наступает самоизлечение. Гименолепидоз сопровождается чередующимися поносами, болями в области живота и истощением. Часто при этом наблюдаются бессонница и эпилептоидные припадки. Для диагностики важно обнаружение яиц в фекалиях. Профилактика сводится к выявлению и лечению больных и соблюдению правил личной гигиены, в первую очередь в детских учреждениях. Имеются сведения, что у мышей и крыс также способны паразитировать некоторые штаммы этого паразита. В связи с этим профилактика гименолепидоза должна включать также борьбу с грызунами. Необходима и постоянная борьба с насекомыми — механическими переносчиками яиц гельминтов.
· Повсеместно
· мучной хрущ (яйцо с онкосферой) — онкосфера, финна типа цистицеркоид
· Человек (финна типа цистицеркоид)
· при поедании теста с хрущами, проглатывание зрелых яиц
Тыквовидный цепень. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
класс Cestoidea
Dipilidum caninum
· Повсеместно
· дипилидоз
· власоеды, личинки блох (яйцо с онкосферой) — онкосфера, финна типа цистицеркоид
· Собаки, лисы, волки, шакалы, иногда – человек (финна типа цистицеркоид)
· проглатывание блох и власоедов при контакте с животными
· тонкий кишечник
- обнаружение зрелых члеников в фекалиях
153.Лентец широкий. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
Тип Plathelmintes
класс Cestoidea
К ним относится несколько видов паразитов из р. Diphyllobothrium, являющихся возбудителями дифиллоботриозов. Гельминты этой экологической группы имеют наиболее архаичный вариант цикла развития:
у них сохранились активно плавающая личиночная стадия — корацидий — и два промежуточных хозяина, обитающие в водной среде, — мелкие планктонные ракообразные из родов Cyclops и Diaptomus, a также рыбы, питающиеся ими. В рачках обитает личинка, называемая процеркоид, в рыбе — плероцеркоид. Примитивной чертой плероцеркоида является его способность к активным перемещениям, причем в случае поедания крупной рыбой более мелких рыб, инвазированных плероцеркоидами, последние пробуравливают стенку кишки и выходят в брюшную полость и мышцы. Таким образом, крупные рыбы на протяжении всей жизни могут накопить в себе сотни плероцеркоидов. Хозяин заражается, поедая инвазированную рыбу. Поэтому основа личной профилактики — термическая обработка рыбных продуктов.
Широкий лентец Diphyllobothrium latum (рис. 20.8, А) — наиболее частый возбудитель дифиллоботриоза у человека. Стробила имеет длину около 10 м. Головка снабжена ботриями — присасывательными щелями, зрелые членики характеризуются маткой розетковидной формы небольших размеров. Матка имеет связь с внешней средой, поэтому созревающие яйца свободно выводятся из нее. Яйца желтоватого цвета, длиной до 71 мкм, имеют на одном полюсе крышечку, а на другом — маленький бугорок. Паразит распространен широко в зоне с умеренным климатом. Заболевание встречается по берегам крупных рек и озер.

Рис. 20.8. Крупные ленточные черви. А — широкий лентец; Б — бычий цепень; В — свиной цепень: а — головки, б — зрелые членики
Жизненный цикл широкого лентеца типичен для этой группы паразитов. Яйца попадают в воду, и из них выходит корацидий, покрытый ресничками. Его проглатывают первые промежуточные хозяева — циклопы. Циклопов с процеркоидами съедают рыбы, в мышцах и икре которых накапливаются плероцеркоиды. Из рыб чаще всего поражаются окуни, ерши, налимы и щуки. В крупных хищных происходит дальнейшее накопление плероцеркоидов. Окончательными хозяевами являются крупные рыбоядные млекопитающие (медведи, собаки и т. д.) и человек. Таким образом, дифиллоботриоз — природно-очаговое заболевание. У человека это заболевание встречается чаще у рыбаков, туристов и людей, употребляющих в пищу слабопросоленную рыбу и икру домашнего производства.
Дифиллоботриоз — это опасное заболевание не только потому, что паразит имеет большие размеры тела и может вызвать кишечную непроходимость, но и в связи с тем, что он находится в антагонистических отношениях с нормальной микрофлорой кишечника, вызывая дисбактериоз. При этом нарушается всасывание витаминов группы В и развивается тяжелая анемия с нарушением кроветворения в красном костном мозге.
Диагностика заболевания основана на обнаружении яиц и обрывков зрелых члеников паразита в фекалиях.
Личная профилактика заражения — см. выше. Общественная профилактика — охрана водоемов от фекального загрязнения.
Кроме этого паразита известны несколько других видов р. Diphyllobothrium, имеющих меньшее значение в связи с малыми ареалами распространения (в Прибайкалье, на территории части Эвенкии, на Сахалине и т. д.) и с тем, что человек ими поражается редко, так как их окончательные хозяева обычно рыбоядные птицы и мелкие млекопитающие. Цикл развития их, методы профилактики и диагностики заболеваний не отличаются от данных, приведенных для широкого лентеца.
Бассейны рек, прибалтика, Дальний Восток
1 — рачки и циклопы (корацидий) — онкосфера – процеркоид, 2 — рыбы (процеркоид) — финна типа плероцеркоид
Рыбоядные млекопитающие, человек (финна типа плероцеркоид)
тонкий кишечник
Эхинококк и альвеококк. Систематическое положение, морфология, географическое распространение, циклы развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики. Отличие личиночных стадий развития.
Тип Plathelmintes
класс Cestoidea
Эхинококк Echinococcus granulosus (рис. 20.9, А) — возбудитель эхинококкоза. Половозрелая форма имеет головку с крючьями и 3—4 членика разной степени зрелости. Последний из них зрелый, он содержит около 800 яиц. Общая длина тела до 5 мм. Яйца по форме и размерам сходны с яйцами свиного и бычьего цепней. Эхинококкоз у человека распространен во всех географических и климатических зонах, преимущественно в регионах с развитым отгонным животноводством.
Жизненный цикл эхинококка связан с хищными животными семейства Псовые (волками, шакалами, собаками), которые являются его окончательными хозяевами. Взрослые членики способны активно ползать, распространяя яйца по шерсти хозяина и в окружающей среде. Их могут проглотить травоядные животные — коровы, овцы, олени или человек, становясь промежуточными хозяевами. Финна эхинококка — пузырь, нередко достигающий 20 см в диаметре. Он заполнен жидкостью с огромным количеством молодых сколексов, постоянно почкующихся от внутренней поверхности стенки финны. Окончательный хозяин заражается, поедая пораженные органы промежуточного.
Растущая финна сдавливает органы, вызывает их атрофию. Постоянное поступление продуктов диссимиляции в организм хозяина вызывает его истощение. Очень опасен разрыв эхинококкового пузыря: жидкость, заключенная в нем, может вызвать токсический шок. При этом мелкие зародышевые сколексы могут распространяться по организму, поражая другие органы. Множественный эхинококкоз обычно заканчивается смертью хозяина.

Рис. 20.9. Ленточные черви, использующие человека как промежуточного хозяина. А — эхинококк; Б — альвеококк: а — половозрелые стадии,
б — финны
Личная профилактика заражения — мытье рук после контактов с пастушьими собаками. Общественная профилактика — обследование и дегельминтизация собак, недопущение скармливания им органов больных животных.
Альвеококк Alveococcus multilocularis (рис. 20.9, Б) — возбудитель альвеококкоза. Половозрелая форма отличается от эхинококка меньшими размерами тела (до 2 мм) и деталями строения крючьев и матки. Финнозная стадия состоит из множества мелких пузырьков, постоянно почкующихся друг от друга наружу, благодаря чему финна представляет собой постоянно растущий узел. В каждом пузырьке находится зародышевая головка паразита. Рост финны разрушает окружающие ткани наподобие злокачественной опухоли. Возможен и отрыв отдельных пузырьков с распространением патологического процесса по организму. Природные очаги альвеококкоза имеются в Сибири, Средней Азии, на Урале и Дальнем Востоке, а также в Северной Америке, в Центральной и Южной Европе.
Жизненный цикл принципиально не отличается от эхинококка, но окончательными хозяевами этого паразита являются дикие хищники — лисы, песцы, волки, реже домашние — собаки, промежуточными — мышевидные грызуны, иногда человек.
Альвеококкоз — более тяжелое заболевание, чем эхинококкоз, в связи с инвазивным характером роста финны.
Личная профилактика — как при эхинококкозе, общественная — соблюдение правил гигиены при обработке шкур промысловых животных, а также запрещение скармливания собакам тушек грызунов.
§
тип Nemathelminthes
класс Nematoda
Toxocara canis и Toxascaris leonina
возбудители токсокароза собак и кошек. Паразиты морфофизиологией и жизненным циклом напоминают аскарид. Миграция личинок в печени вызывает гепатит, а в головном мозге — менингитоподобные явления и даже смерть. Часто заражаются дети, активно контактирующие с домашними животными.
Профилактика — гигиеническое содержание домашних животных.
· геогельминт
· распространен повсеместно
· яйцо с развитой личинкой
· кишечник — кровеносные сосуды — ткань печени, легких, гм, лимфатических узлов, глаза
· заглатывание яиц с пищевыми продуктами, при контакте с шерстью собак
· локализация: печень, легкие, гм, органы зрения, лимфатические узлы
· профилактика: соблюдение правил личной гигиены, дегельминтизация собак
158.Власоглав человеческий. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Nemathelminthes
класс Nematoda
Trichocephalus trichiurus
· геогельминт
· Передний конец тела этого червя вытянут. В нем находится только пищевод. Задняя часть тела утолщена, здесь располагаются кишка и половая система. Общая длина паразита 3—5 см. Яйца светлые овальные, прозрачные, с двумя характерными пробочками, длиной до 50 мм.
· Очаговое распространение
· Инвазионная стадия: яйко с развитой личинкой
· Не мигрирует
· Заражение: проглатывание яиц с грязными овощами и фруктами
· Локализация: слепая кишка, червеобразный отросток
· Профилактика: мыть руки, овощи и фрукты
· Власоглав поселяется в слепой и восходящей части толстой кишки, прикрепляясь к стенке передним концом тела и питаясь кровью и тканевой жидкостью из глубоких слоев слизистой оболочки. Яйца, выводимые с фекалиями, в почве достигают инвазионности за 3—4 недели. Заражение человека осуществляется при проглатывании инвазионных яиц. Половой зрелости паразиты достигают в кишке уже через несколько недель. Патогенное действие власоглава связано с интоксикацией хозяина продуктами жизнедеятельности и с нарушением функции кишечника. При массивной инвазии, возможны местные воспалительные явления, а также головные боли, головокружение и даже судороги.
· В связи с тем что власоглав не питается содержимым кишечника и плотно прикреплен к его стенке, выведение этого паразита из организма человека более сложно по сравнению с многими другими геогельминтами.
159.Острица детская. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, методы лабораторной диагностики, меры профилактики. Обоснование методов без медикаментозного лечения.
тип Nemathelminthes
класс Nematoda
Enterobius vermicularis
· возбудитель энтеробиоза — самого распространенного гельминтоза, который встречается в основном у детей.
· Мелкий червь с выраженным половым диморфизмом: самка достигает в длину 12 мм, тело ее прямое, сзади заостренное. Самцы мельче (до 5 мм) и свернуты спирально. Яйца овальны, несколько асимметричны, прозрачны и бесцветны, длиной до 50 мкм.
· Жизненный цикл в целом соответствует циклу развития власоглава, но яйца достигают инвазионности после откладывания не за несколько недель, а всего за несколько часов. Поэтому при энтеробиозе часто происходит аутоинвазия. Обитают острицы в нижнем отделе тонкой кишки, питаясь ее содержимым. Самки со зрелыми яйцами выползают из анального отверстия и откладывают яйца на коже промежности, вызывая при этом сильный зуд. При расчесывании зудящих мест яйца попадают на руки, затем на белье и игрушки. При проглатывании яиц из них быстро развиваются взрослые паразиты, продолжительность жизни которых около 1 мес. Патогенное действие остриц связано с тем, что зуд промежности приводит к нарушению сна, к нервному истощению. Возможно также нарушение стенки кишечника.
· Лабораторная диагностика имеет особенности: в фекалиях яйца обнаруживаются редко, поэтому их исследуют в мазке с перианальных складок промежности.
· Личная профилактика — соблюдение правил личной гигиены, особенно тщательное мытье рук после сна, короткая стрижка ногтей. Общественная — регулярно обследовать детей в детских учреждениях, лечение больных, регулярные обследования также работников детских учреждений и предприятий общественного питания.
· Геогельминт
· повсеместно
· инвазионная стадия: яйцо с развитой личинкой
· не мигрирует
· нижний отдел тонкого кишечника, верхний отдел толстого
160.Анкилостомиды. Систематическое положение, морфология, географическое распространение, циклы развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики. Очаги анкилостомидозов и пути их ликвидации.
тип Nemathelminthes
класс Nematoda
семейство Анкилостомиды
Кривоголовка двенадцатиперстной кишки Ankylostoma duodenale и некатор Necator americanus
Отличаются друг от друга формами ротовой капсулы и особенностями географического распространения. Кривоголовка имеет в ротовой полости четыре хитиновых зуба, а некатор — две широкие режущие пластинки полулунной формы. Оба паразита широко расселены по всему тропическому и субтропическому поясу, однако некатор чаще встречается в зонах с более жарким климатом. В остальном оба вида сходны настолько, что даже заболевания, вызываемые ими, не дифференцируются и называются анкилостомидозами. Размеры анкилостомид около 10 мм. Головной конец загнут на брюшную сторону; яйца овальные, прозрачные, длиной до 60 мкм.
Жизненный цикл анкилостомид своеобразен и свидетельствует о тесной связи их со свободноживущими предковыми формами. Яйца, попадающие в почву, быстро развиваются, и из них вскоре выходят личинки, которые, дважды линяя, через несколько месяцев становятся инвазионными и могут попадать в организм человека либо с загрязненными почвой овощами и фруктами, либо за счет активного внедрения через кожу.
Эта особенность обеспечивает возможность существования подземных очагов анкилостомидозов в шахтах, находящихся в зонах умеренного пояса. Основным условием возникновения таких очагов является высокая влажность грунта и загрязненность его фекалиями. Попав в кровь, личинки проделывают по сосудам путь, характерный для всех видов гельминтов этой группы. Окончательная локализация — двенадцатиперстная кишка, к ворсинкам которой они прикрепляются ротовыми капсулами, повреждая их и питаясь кровью и клетками слизистой оболочки. Оба паразита выделяют в ранки антикоагулянтные вещества и могут вызвать кишечные кровотечения и аллергизацию больных.
Лабораторная диагностика — как при всех геогельминтозах.
Профилактика — кроме общих мер, описанных выше, обязательное ношение обуви в районах, где распространены эти заболевания.
· Геогельминт
· тропический и субтропический пояс
· инвазионная стадия: филляритивная личинка после 2ой линьки
· Миграция: кожа — кровеносные сосуды — сердце — легочные артерии — капилляры альвеол — глотка — ктшечник
· тонкий кишечник — 12перстная
161.Угрица кишечная. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Nemathelminthes
класс Nematoda
Strongyloides stercoralis
возбудитель стронгилоидоза. Это очень мелкий гельминт, до 2,2 мм длиной. Из всех известных паразитов человека — наименее специализированный.
Жизненный цикл этого червя характеризуется значительной вариабельностью. Он может проходить целиком во внешней среде либо частично или полностью в организме хозяина. Чаще встречается чередование свободноживущего и паразитического поколений.
Паразитические половозрелые угрицы живут в кишечнике человека. Из яиц уже в кишечнике выходят личинки, судьба которых может быть двоякой. В первом случае после линьки в почве они могут стать инвазионными и проникают в организм человека либо через кожу, либо перорально с фруктами и овощами. Дальнейшее развитие в этом случае проходит так же, как у анкилостомид. Во втором случае личинки могут достичь половозрелости в почве.
Взрослые самки откладывают яйца и дают начало свободноживущим поколениям. При изменении условий жизни личинки могут стать инвазионными для человека и вновь перейти к паразитическому существованию. Присутствие паразитов в кишечнике вызывает чередующиеся поносы и запоры. Часто при этом паразиты заселяют и жёлчные ходы печени, вызывая желтуху.
Аллергизация организма при стронгилоидозе очень сильна, особенно на миграционной стадии развития.
Описаны случаи течения стронгилоидоза в течение 25 лет. Это объясняется не только длительностью жизни этого паразита,но ивозможностью аутоинвазии.
Как и анкилостомиды, угрица кишечная встречается в основном в тропической и субтропической зонах. Особенно широко расселена в Юго-Восточной Азии, Южной и Центральной Америке. Возможно существование подземных очагов в зонах с умеренным климатом, как и при анкилостомидозах.
При диагностике надежным признаком заболевания является обнаружение живых личинок в фекалиях, рвотных массах и при дуоденальном зондировании.
Профилактика стронгилоидоза соответствует профилактике анкилостомидозов.
162.Трихинелла спиральная. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Nemathelminthes
класс Nematoda
Trichinella spiralis
возбудитель трихинеллеза— мелкий гельминт длиной до 4 мм. Распространен очень широко, на всех континентах и во всех природно-климатических зонах. Этому благоприятствуют такие особенности биологии паразита, как способность личинок переживать неблагоприятные условия на протяжении десятков лет. Хозяевами трихинеллы могут быть различные хищные и всеядные млекопитающие, а также человек. Распространение трихинелл происходит обычно при поедании животными друг друга.
Человек заражается, поедая мясо зараженных животных, чаще всего свиней. Проглоченные личинки в кишечнике быстро достигают половой зрелости. Оплодотворенные самки рождают живых личинок, которые пробуравливаются через стенку кишечника и, транспортируясь кровью, оседают в поперечно-полосатых мышцах: чаще всего в диафрагме, межреберных, дельтовидных. Здесь они после разрушения части мышечных волокон спирально скручиваются и инкапсулируются.
При попадании трихинелл в ослабленный организм возможно развитие первого поколения личинок не в мышцах хозяина, а в ворсинках его кишечника, после разрушения которых личинки вновь возвращаются в просвет кишки, достигают там половой зрелости и размножаются. Следующее поколение личинок уже оседает в мышцах хозяина. Этим достигается резкое увеличение размеров популяции паразита, которые в обычных случаях размножаются у одного хозяина лишь один раз, и соответственно резкое утяжеление заболевания, часто приводящее к смерти.
Личинки трихинелл после гибели хозяина сохраняют жизнеспособность даже после разложения его трупа. В пищеварительной системе трупоядных наземных и даже водных животных — жуков-мертвоедов, почвенных червей, мелких ракообразных, рыб, хищных птиц и чаек — личинки сохраняют жизнеспособность в течение нескольких дней. Перечисленные животные выступают в цикле развития паразитов как транспортные хозяева и способны передавать возбудителя по цепи питания до тех пор, пока личинки не попадут в организм хозяина, в котором возможно нормальное развитие паразита. Сложными цепями питания с передачей личинок транспортными хозяевами обеспечивается фактически всесветное распространение трихинеллы и существование таких природных очагов трихинеллеза, как очаги даже в зоне арктических пустынь.
Известно, что 67% ездовых собак в Гренландии и 50% белых медведей о. Шпицберген, многие тюлени и даже некоторые китообразные поражены трихинеллезом. Основной источник питания этих животных — рыба, морские беспозвоночные, яйца и птенцы морских птиц и т. д. Эти животные становятся транспортными хозяевами трихинеллы после того, как разлагающиеся трупы и помет птиц оказываются в воде и личинки паразита попадают в организм водных ракообразных, которыми питаются более крупные беспозвоночные и рыбы. Поедая их, млекопитающие постепенно накапливают в организме огромное количество трихинелл, становясь опасным источником заражения человека. Свиньи как наиболее частый источник инвазий человека заражаются, поедая не только крыс и мышей, но и насекомых, червей, рыбу, трупы и помет птиц, в которых сохраняются жизнеспособные личинки.
Патогенное действие трихинеллы включает как общеаллергические явления, так и нарушение функций мышц.
Диагноз основывается на данных анамнеза — употребление мяса диких животных и не проверенной ветеринарной службой свинины, а также на результатах биопсии мышц.
Личная профилактика — тщательная термическая обработка свинины и особенно мяса диких животных. Общественная профилактика — санитарный надзор в свиноводстве, проверка свинины на торговых точках и на предприятиях общественного питания.
Трихинеллез – природно-очаговое заболевание. Половозрелые особи обитают в кишечники человека. Самки достигают размера 4 мм, самцы – 2 мм. Природные резервуары – лисы, курицы, кабаны, крысы, мыши, свиньи и т.п. при поедании зараженного мяса происходит инвазирование. Через 3 суток достигают размера взрослых особей. Живет 2 месяц, производит до 2000 личинок. Имеет место живорождение. Личинки часто оседают в поперечно-полосатых мышцах. Симптомы заболевания часто сходны с брюшным тифом. Диагноз ставится при изучении инкапсулированных личинок в биоптатах.
· Биогельминт
· повсеместно кроме Австралии
· инвазионная стадия: инкапсулированная личинка
· кишечник — лимф система — кровеносная система — специальные группы мышц
· взрослые особи — в тонком кишечнике, личинки — в жевательных, межреберных, дельтовидных мышцах, диафрагме
· сан-гигиен контроль на скотобойнях и рынках
163.Ришта. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Nemathelminthes
класс Nematoda
Dracunculus medinensis
возбудитель дракункулеза. Длина самки до 120 см, самца — только 2 см. Заболевание распространено в зонах с тропическим и субтропическим климатом, раньше встречалось в Средней Азии.
Жизненный цикл ришты связан с водной средой. Окончательные хозяева ришты — человек, обезьяны, домашние и дикие млекопитающие, у которых черви локализуются под кожей конечностей. У человека наиболее частая локализация — под кожей ног в области суставов. Описаны случаи обнаружения гельминта под серозной оболочкой желудка, под мозговыми оболочками, в стенке пищевода. Над передним концом зрелой самки образуется кожный пузырь, заполненный серозной жидкостью. Человек при этом ощущает сильный зуд, проходящий при соприкосновении с водой. Опускание ног в воду сопровождается разрывом пузыря и рождением живых микроскопических личинок, которые сразу проглатываются промежуточными хозяевами — циклопами. В полости тела циклопов они через несколько дней достигают инвазионности и при проглатывании с водой таких рачков попадают в кишечник, а затем мигрируют под кожу.
Весь жизненный цикл ришты длится 1 год. Интересно, что развитие паразитов у инвазированных людей происходит синхронно, таким образом, что самки становятся способными рождать личинок одновременно почти у всех носителей паразита. Этим достигается резкое повышение вероятности заражения огромного количества циклопов, а затем и основных хозяев в течение небольшого интервала времени. Эта особенность жизненного цикла ришты имеет огромное адаптивное значение в зонах с засушливым климатом и редкими, повторяющимися из года в год дождевыми периодами. Она имеет и существенное медицинское значение в связи с тем, что в очагах распространения дракункулеза выявляется большое количество пораженных этим паразитом людей в течение небольшого временного интервала. Это облегчает врачу постановку диагноза, лечение и проведение профилактических мероприятий.
Кроме общих аллергических реакций обязательно проявляется и местное воздействие паразита: локальные воспалительные реакции и нарушение функций суставов, прилежащих к зоне поражения.
Диагностика при типичной локализации проста: паразит виден под кожей. Атипичное расположение гельминта требует применения иммунологических реакций.
Личная профилактика также проста — кипячение или фильтрация питьевой воды, взятой из открытых водоемов. Общественная профилактика — современное водоснабжение обеззараженной водой; выявление и лечение больных гарантирует успех в борьбе с этим заболеванием.
164.Филярии. Систематическое положение, морфология, географическое распространение, цикл развития, пути заражения, патогенное действие, обоснование методов лабораторной диагностики, меры профилактики.
тип Nemathelminthes
класс Nematoda
сем. Filarioidea
Трансмиссивный способ передачи вообще очень редок среди гельминтов. Он характерен только для этой небольшой группы нематод. Все паразиты этой группы распространены в тропиках. Они относятся к сем. Filarioidea и вызывают заболевания — филяриатозы. Половозрелые особи, или филярии, обитают в разных тканях внутренней среды основного хозяина, а рождаемые ими личинки, или микрофилярии, периодически поступают в кровь и лимфу и некоторое время находятся там (рис. 20.18). Если в этот момент на хозяина нападают кровососущие насекомые, микрофилярии с кровью поступают в желудок, оттуда в мышцы переносчика и за одну или несколько недель достигают там инвазионности, переходят в хоботок переносчика и затем, при укусе, — в кровь основного хозяина. У некоторых видов филярии может быть несколько основных хозяев (человек, обезьяна, собака и т. д.), а переносчики всегда специфичны.
В связи с тем что в переносчиках происходит развитие личинок паразита, они являются одновременно и промежуточными хозяевами. Длительные взаимные адаптации филярий, их хозяев и переносчиков привели к своеобразному эффекту — у большинства филярий происходит выход личинок в кровяное русло в строго определенные часы суток, соответствующие периодам максимальной активности переносчиков. Так, у филярий, переносчиками которых являются комары, личинки поступают в кровь в вечерние и ночные часы суток. Если переносчиками являются слепни, то появление микрофилярий в крови приурочено к утренним и дневным часам. В том случае, когда переносчиками служат мокрецы, активность которых больше зависит от влажности воздуха, выход микрофилярий в кровь лишен периодичности. Эти особенности биологии филярий необходимо учитывать при постановке диагноза и брать кровь у больных для обнаружения в ней микрофилярий в то время, когда наличие их там наиболее вероятно. Профилактика филяриатозов — выявление и лечение больных, борьба с переносчиками.
§
| Название паразита и болезни | Географическое распространение | Промежу-точный хозяин и переносчик | Основной хозяин | Локализация половозрелых форм, симптомы | Время появления микрофилярий в крови |
| Wuchereria banctofti Вухерериоз Brughiamalayi Бругиоз | Тропическая Азия, Африка, Южная Америка Южная и Восточная Азия | Комары » | Человек, высшие обезьяны Человек, обезьяна, кошачьи | Лимфатические узлы и сосуды Застой лимфы, отеки и развитие слоновости,аллергия То же | Вечер и ночь Ночь |
| Onchocerca volvulus Онхоцеркоз | Эваториальная Африка, Центральная, Северная и Южная Америка | Мошки | Человек | Под кожей груди, конечностей, головы Образование узлов. Слепота при поражении глаз, аллергизация | Нет периодичности |
165.Методы овогельминтоскопии 166.ТИП ЧЛЕНИСТОНОГИЕ. Классификация. Характерные черты организации. Медицинское значение.
Медицинская арахноэнтомология изучает представителей типа Членистоногие Arthropoda, имеющих медицинское значение. Тип Членистоногие содержит более 1,5 млн. видов. Он характеризуется наличием хитинового покрова — скелетного и защитного образования — и членистых конечностей. Тело состоит из сегментов, сливающихся в три отдела: голову, грудь и брюшко. В некоторых группах членистоногих голова и грудь представляют собой единое образование — головогрудь, иногда тело вообще не расчленено. На голове расположены органы чувств и ротовой аппарат — видоизмененные конечности. В пищеварительной системе имеются сложные железы. Органы дыхания в зависимости от систематического положения и образа жизни жабры, мешковидные легкие или трахеи. Кровеносная система незамкнутая, сердце находится на спинной стороне. На брюшной стороне — нервная цепочка из частично слившихся ганглиев, среди которых самые крупные — подглоточный и надглоточный — расположены на переднем конце тела.
Наибольшее медицинское значение имеют классы Паукообразные и Насекомые. В этих классах встречаются временные и постоянные паразиты, переносчики и возбудители инфекционных и паразитарных заболеваний. В классе Ракообразные встречаются только отдельные виды — промежуточные хозяева некоторых гельминтов.
Членистоногие – самый многочисленный тип.
Характерные признаки:
1) двусторонняя симметрия;
2) трехслойная организация;
3) гетерономная метамерия;
4) слияние сегментов тела в отделы;
5) членистые конечности;
6) обособление мышц и появление поперечно-полосатых мышц;
7) наружный хитиновый скелет;
8) наличие полости тела (миксонель);
9) наличие систем органов (пищеварительная, дыхательная, выделительная, кровеносная, нервная, эндокринная, половая).
Пищеварительная система.
Состоит из 3х отделов: переднего, среднего и заднего. Средний отдел имеет пищеварительные железы, задний оканчивается анальным отверстием.
Дыхательная система.
В зависимости от образа жизни, в водной среде – жабры, на суше – трахея и легкие. У мелких насекомых газообмен осуществляется всей поверхностью тела.
Выделительная система.
Представлена мальпигиевыми сосудами. Располагается на границе средней и задней кишки. Есть специальные выросты. У некоторых сохраняются пронефридии.
Кровеносная система.
Трубчатое сердце на спинной стороне тела. Кровеносная система незамкнутого типа.
Нервная система.
ЦНС – головной мозг и брюшная нервная цепь.
Функцию регуляции выполняет также и эндокринная система. Голова несет глаза, усики и др.
Половая система.
Выражен половой диморфизм. Развитие прямое или с превращением.
167.Класс Паукообразные. Классификация. Характерные черты организации. Медицинское значение.
Прогрессивные черты паукообразных.
1. гетерономная метамерия;
2. появление конечностей;
3. развитие мышечной системы;
4. наружный хитиновый скелет;
5. появление сердца;
6. усложнение в строении нервной системы;
7. усложнение органов чувств.
Есть 5 подтипов паукообразных. Особое медицинское значение у 3 из них:
– Жабродышащие (Branchiata) – раки, крабы, циклопы.
– Хелицеровые (Chelicerata).
– Трахейнодышащие (Tracheata) – как и предыдущий подтип – паразиты, ядовитые формы, резервуары и переносчики.
TRACHEATA
Трахейнодышащие.
Arachnoidea (паукообразные) – 5000 видов. Имеют все приспособления к жизни на суше. Характерно слияние сегментов тела, члеников: головогрудь, брюшко. Интересны отряды скорпионы, пауки и клещи.
Гнатосома включает ротовые органы.
Отдельные участки собственно тела уплотняются, образуются диски, щитки. На передней поверхности идиосомы могут быть глаза. Органы осязания – щупики, или пальпы, состоят из 4 члеников.
Имаго имеет 4 пары конечностей. Нога состоит из 5-7 члеников, последний членик – лапка – имеет подушечки и коготки.
Дыхание – трахейное. Личинки и мелкие особи осуществляют дыхание всей поверхностью тела.
Характерен половой диморфизм. Раздельнополые организмы. Самцы меньше самок. У самок только частичная спинная хитинизация, у самцов – сплошная. Кровососущие только самки, самцы же питаются соком растений. Хитиновый покров мешает увеличению размеров тела.
Постоянные и временные паразиты, частые переносчики трансмиссивных заболеваний.
168.Клещи. Систематическое положение, морфология, географическое распространение, циклы развития. Медицинское значение клещей семейств Иксодовые Ixodidae (собачий, таёжный, пастбищный, хиалома), Аргазовые Argasidae (поселковый), Акариформные Acariformes (чесоточный); надсемейства Гамазовые (крысиный); подотряда Trombilidiformes (семейства Краснотелковые Trombiculidae и Demodicidae (железница угревая). Домашний клещ.
Иксодовые клещи довольно крупные — от нескольких миллиметров до 2 см в зависимости от степени насыщения. На переднем конце тела ротовой аппарат сильно выступает вперед. Основным компонентом хоботка является гипостом — длинный уплощенный вырост, несущий направленные кзади острые зубцы. Хелицеры имеют вид колющих стилетов, зазубренных с латеральных сторон. С их помощью в коже хозяина образуется резаная ранка, и в нее вводится гипостом.
Первая порция слюны, вводимая в ранку при укусе, обладает способностью застывать вокруг хоботка. В результате разведенные в стороны хелицеры и гипостом прочно заякориваются. Так достигается надежное прикрепление клеща к хозяину, позволяющее ему питаться на протяжении от нескольких часов до 15—20 дней. Средняя кишка имеет многочисленные выросты, заполняющиеся кровью при питании. Здесь кровь может сохраняться до нескольких лет (рис. 21.1). Вся спинная сторона самца покрыта нерастяжимым хитиновым щитком, у самки такой щиток занимает не более половины поверхности тела, поэтому покровы самки значительно более растяжимы и она может выпить гораздо больше крови. После питания самки откладывают от 1500 до 20 000 яиц в лесную подстилку, трещины почвы, в норы грызунов. Личинки имеют три пары ходильных ног. Они питаются кровью ящериц и мелких грызунов.
Следующая стадия жизненного цикла — нимфа.
Она значительно крупнее личинок, имеет три пары ног и питается на зайцах, белках, крысах. После линьки нимфа превращается в половозрелую стадию.
Взрослый клещ сосет кровь крупных домашних и диких копытных, лис, собак и человека. В связи с тем что для каждого периода при переходе к следующей стадии цикла развития клещам необходимо питаться один раз, большинство иксодовых клещей меняет трех хозяев. Такой цикл развития называют треххозяинным. Многие иксодовые клещи пассивно подстерегают своих хозяев, однако они локализуются в таких местах, где встреча с прокормителем наиболее вероятна: обычно они располагаются на высоте до 1 м на концах веточек и листьев кустарников вблизи троп, по которым передвигаются животные. Некоторые виды совершают активные поисковые движения.
Иксодовые клещи переносят возбудителей опасных заболеваний. Среди этих заболеваний наиболее известен клещевой весенне-летний энцефалит. Вирусы размножаются в организме клеща и накапливаются в слюнных железах и яичниках. При кровососании происходит транс-миссивная передача вирусов хозяину-прокормителю, а при откладке яиц — трансовариалъная передача следующему поколению клещей. Наибольшее значение как переносчики и резервуары возбудителей имеют следующие виды иксодовых клещей.
IXODIDA
Все – облигатные гематофаги. Сезонные эктопаразиты. Активны в период от ранней весны до поздней осени. Зимой впадают в состояние анабиоза. Поскольку у них не кровь, а лимфа, она не замерзает. Характерно многодневное питание, длящееся 15-20 дней. Для этого клещ фиксируется на теле хозяина. У него есть гепастом (пластинка с направленными к телу клеща зубцами, внутри – канал), хелицеры. Когда клещ попадает на кожу, выделяется первая порция слюны. После прикрепления – вторая: гидролитические ферменты, антикоагулянты. Образуется очаг некроза, формируется полость, заполненная тканевой жидкостью и кровью, а также экссудатом. Лимфой питается имаго. К концу кровососания самки увеличиваются в 200 раз в весе и в 4-6 раз в размере. Сытые самки отпадают на землю и через 4, 12, 60 дней откладывают яйца в умеренно увлажненные и затемненные места. Количество яиц от 100 до 10000. после откладывания яиц самка гибнет. Эмбриональное развитие длится 2-4 недели. Личинка мелкая – 0,6 – 0,38 мм, не имеет половых органов, но есть идиосома, гнатосома. 3 пары конечностей. Личинки голодные, ищут хозяина. 2-4 дня питаются кровью на мелких млекопитающих, затем происходит линька. Возникает 8-ногая нимфа, размеры которой близки к размерам имаго, но половых органов нет. Нимфы питаются кровью 3-5 дней. Затем линяют и превращаются в имаго. Длительность развития зависит от вида клещей, от возможности отыскания хозяина и от условий внешней среды.
Живут в лесах, зарослях кустарников, лугах, пустошах.
В России известно около 70 видов и подвидов. Каждый вид имеет границы распространения.
Наиболее часто встречаются:
Ixodes persulcatus
Ixodes ricinus
Dermacentor marginatus
Dermacentor pictus
Общая продолжительность жизни 3-6 лет. Длительность голодания имаго 2-3 года. Подразделяются на
– однохозяйные – личинки, нимфы и имаго обитают на одном хозяине.
– двуххозяйные – голодная личинка и нимфа – на одном хозяине, а имаго ищет нового.
– треххозяйные – все стадии питаются и обитают на разных хозяевах.
Таежный клеш, Ixodes persulcatus. Размеры самца 2,5 мм, самки — до 4 мм. Распространен в таежной зоне Евразии от Дальнего Востока до горных районов Центральной Европы. Живет до 3 лет. Этот вид клещей особенно опасен, так как они наиболее часто нападают на человека.
Ixodes persulcatus
Самки – 3 мм, самцы – 2,5.
Агрессивен в отношении человека (активнее собачьего). Цикл развития – 2-3 года. Обитает в смешанных лесах от Балтийского моря до Дальнего Востока, Камчатки, Курильских островов.
Собачий клещ Ixodes ricinus (рис. 21.2). Встречается часто в смешанных, лиственных лесах и кустарниковых зарослях большей части Евразии. От таежного клеща отличается некоторыми деталями строения и более длительным циклом развития — до 7 лет.
Ixodes ricinus
Самки – 3мм, самцы – 2,5.
Личинки и нимфы обитают на грызунах, птицах, ящерицах. Взрослые клещи – на крупных и средних диких и домашних животных. Зайцы и ежи – прокормители всех фаз. Цикл развития составляет 3-4 года. Клещ распространен в смешанных лесах, кустарниковых зарослях. От западной границы до Волги, от Карелии до юга, а также в Северной Америке и Западной Европе.
Пастбищный клещ Dermacentor sp. (рис. 21.2). Чаще встречается на лугах, пастбищах, в лесостепной зоне и в горных лесах. От клещей рода Ixodes отличается более коротким ротовым аппаратом, беловатым эмалевым рисунком на поверхности спинного щитка и фестончатым, а не гладким задним краем тела.
Dermacentor marginatus et pictus
На спинном щитке у взрослых особей имеются белые эмалевые рисунки на коричневом фоне. Имаго с конца марта и до начала июня паразитирует на мелких и средних диких и домашних животных. Личинки и нимфы обитают на грызунах. Цикл развития короткий – около года. Встречаются на лугах, кустарниковых зарослях, лесных луговинах. Распространены от западной границы до Западной Сибири, в Казахстане и др.
Клещи рода Hyalomma (рис. 21.2) — крупные иксодовые клещи (более 5 мм). Обитают в степной зоне и в горах субтропических районов Южной Европы. Характерной чертой строения являются очень толстые и длинные ноги.
Аргазовые клещи отличаются от иксодовых тем, что их ротовой аппарат располагается на вентральной стороне тела и совершенно не выступает вперед. Щитка на спинной стороне тела нет. Вместо него покровы снабжены многочисленными хитиновыми бугорками и бляшками и очень сильно растяжимы. По краю тела со всех сторон проходит широкий рант. Продолжительность жизни более 20 лет. Распространены в странах с теплым или жарким климатом. Часто встречаются в Закавказье и Средней Азии, причем не только в естественных биотопах, но и в антропогенных ландшафтах и даже в хозяйственных постройках и жилищах человека.
АРГАЗОВЫЕ.
Тело удлиненное, овально-яйцевидное, мешковидное. Подушечки есть только у личинок. Нимфальных стадий много 2-6. Облигатные кровососы. Живут в норах, глинобитных строениях, камнях и других закрытых местах. Нападают на добычу ночью, в затемненных местах – и днем.
Ornithodorus papillitus (поселковый клещ).
Самка – 6-10 мм, самец – 3-7.
Тело окаймлено валиком, овально-яйцевидная форма, темно-серая окраска. Распространен в районах, близких к Узбекистану, Таджикистану. Обитают в пещерах, норах, временных укрытиях черепах ежей, в поселках – в глинобитных постройках. Питается кровью любых животных. Длительность кровососания – не более часа. В эксперименте с содержанием голодных и сытых самок наблюдался каннибализм.
Размножаются в июле-сентябре. Самка многократно откладывает яйца с годичным интервалом. Может голодать 20-25 лет, личинки – 0,6 лет, нимфы – до 10 лет. В клеще сохраняются спирохеты – возбудители клещевого возвратного тифа.
В большинстве случаев нет строгой специфичности.
Среднеазиатский возвратный тиф вызывает спирохета Borrelia sogdiana, ее переносчиком является поселковый клещ Omithodorus papillipes (рис. 21.3). Болезнь характеризуется периодическими лихорадочными приступами и воспалительными явлениями в дыхательной системе, а также поражениями центральной нервной системы. Обычно заболевание заканчивается выздоровлением через 2,5—4 мес. с момента заражения.
Надсемейство Гамазовые клещи Gamasoidea — очень мелкие клещи, размером от 0,2 до 2,5 мм. Ведут хищнический или эктопаразитический образ жизни. Некоторые виды даже приспособились к эндопаразитиз-му. Все тело покрыто длинными щетинками. Ноги длинные и очень хорошо развиты. Хелицеры у паразитических видов превращены в колющие стилеты. Множественные укусы паразитических гамазовых клещей вызывают у человека местные поражения кожи — дерматиты. Ряд клещей этой группы имеют значение как переносчики возбудителей некоторых вирусных, риккетсиозных и бактериальных заболеваний. Наиболее известны среди них крысиный сыпной тиф, вирус которого передается клещом Omithonyssus bacoti, риккетсиоз Q — лихорадка, передающаяся куриным клещом Dermanyssus gallinae (рис. 21.4 А, Б), а также туляремия, в переносе которой участвуют гамазовые клещи, паразитирующие на водяных крысах.
Семейство Краснотелковые клещи Trombiculidae включает ряд видов, имеющих медицинское значение. Характерным для этих видов является личиночный паразитизм: половозрелые стадии цикла развития протекают в почве, личинки же — временные кровососущие эктопаразиты человека и животных. Большинство из них имеет ярко-красные бархатистые покровы (рис. 21. 4, В). Укусы личинок вызывают образование волдырей и сильный зуд. Массовое нападение личинок на человека наблюдается во второй половине лета, во время полевых работ. При этом проявления дерматита могут быть очень сильными. Это состояние называют осенней эритемой. В результате расчесывания волдырей возможно внесение в кожу бактерий, осложняющих течение болезни.
Некоторые краснотелки Дальневосточного региона распространяют тяжелый трансмиссивный риккетсиоз — японскую речную лихорадку, или цуцугамуши. Природным резервуаром этого заболевания являются дикие грызуны.
Особый интерес представляют в настоящее время так называемые домашние клещи, обитающие в матрацах, коврах, мягкой мебели и в постельном белье. Наиболее известный домашний клещ — Dermatophagoides pteronyssinus (рис. 21.5). Размеры его около 0,1 мм, а в 1 г домашней пыли обнаруживается от 100 до 500 экземпляров клещей этого вида. Подсчитано, что в матрасе одной двуспальной кровати может одновременно обитать популяция, состоящая из 2 млн. особей этого вида. Исследования аллергологов последних лет показали, что аллергические реакции к антигенам этого клеща обнаруживаются у 45—85% страдающих бронхиальной астмой, в то время как у неастматиков аллергия к клещам встречается в 5—30% случаев.
В течение нескольких последних лет в Папуа — Новая Гвинея вместе с мягкой мебелью и постельными принадлежностями из Европы были занесены также и домовые клещи. Заболеваемость бронхиальной астмой среди местного населения при этом возросла в 50 раз. Исходя из того что в настоящее время в развитых странах до 10% детей страдают бронхиальной астмой, причем для большинства из них аллергенами являются компоненты домашней пыли, становится понятным медицинское значение клещей этой экологической группы.
Основные меры борьбы с клещами, обитающими в пищевых продуктах,— понижение влажности и температуры в помещениях, где они хранятся.
Борьба с домовыми клещами состоит в первую очередь в частых влажных уборках помещений, использовании мебели, подушек и матрацев из синтетических материалов, в которых эти животные не могут поселяться.
Железница угревая Demodex folliculorum (рис. 21.6, Б) — возбудитель демодикоза. Эти клещи имеют червеобразную форму, длиной не более 0,4 мм. Они обитают в сальных железах и волосяных фолликулах кожи лица, шеи и плеч, располагаясь головным концом вниз, обычно группами по четыре особи. Часто встречаются у совершенно здоровых лиц, не вызывая никаких симптомов. Однако у ослабленных людей, в особенности со склонностью к аллергическим реакциям, железницы могут активно размножаться, вызывая закупорку протоков сальных желез, возникновение угрей розового цвета с гнойным содержимым.
Для диагностики демодикоза выдавленное содержимое сальной железы или выдернутую ресницу помещают в каплю бензина на предметное стекло и микроскопируют. Таким образом могут быть обнаружены взрослые формы, нимфы, личинки и яйца паразита.
Расселение угрицы по человеческим популяциям происходит при личном контакте и пользовании общими полотенцами и бельем, причем у 40—60% населения можно обнаружить угрицу, живущую как комменсал. Поэтому профилактика демодикоза сводится большей частью к лечению основных заболеваний, ослабляющих организм, а также выявлению и лечению больных с выраженными аллергическими реакциями.
Sarcoptes scabiei — возбудитель чесотки человека. Близкие виды вызывают чесотку у домашних и диких животных, но строгой специфичностью по отношению к хозяину не обладают. Человека могут поражать чесоточные зудни лошадей, овец, коз, верблюдов, собак и других животных. Они вызывают характерные поражения кожи, но на человеке живут недолго и почти не размножаются.
Самка чесоточного зудня длиной до 0,4 мм. Самец значительно меньше. Все тело покрыто щетинками разной длины, и на конечностях имеются присоски. Ротовой аппарат адаптирован к прогрызанию ходов в толще эпидермиса, где обитает этот паразит. Самка откладывает до 50 яиц за всю жизнь, которая длится около 15 сут. Чесоточные зудни могут поселяться в любом месте кожи, но наиболее часто — на нежных ее частях: в межпальцевых промежутках, на сгибах рук, на половых органах. Перемещаясь в толще кожи, клещи раздражают нервные окончания и вызывают нестерпимый зуд, усиливающийся ночью. При расчесывании ходов клещей они переносятся на другие участки кожи или на другого человека. Так происходит расселение клещей по телу хозяина и заражение чесоткой здоровых людей. Заразиться чесоткой можно также и при пользовании одеждой, постельными принадлежностями и личными вещами больного.
Диагностика чесотки проста, так как поражения кожи клещами очень характерны. Они представляют собой прямые или извилистые полоски грязно-белого цвета. На одном из концов хода располагается пузырек, в котором находится клещ. Его можно перенести на предметное стекло в каплю 50 %-ного раствора глицерина и микроскопировать.
Для профилактики чесотки необходимо соблюдение правил личной гигиены, выявление и лечение больных и дезинфекция их одежды, белья и полотенец, а также осторожность при общении с животными.
Влияние клещей на организм человека и других животных.
при нарушении целостности кожных покровов создает ворота для проникновения возбудителей;
токсичная слюна – местная кожная реакция. При массовом нападении – лихорадка, нервные болезни;
аргазовые вызывают клещевые параличи, приводящие к гибели молодых животных;
большая потеря крови 100 самок Dermacentor marginatus – 450 мл.
слюна обладает антигенными свойствами, поэтому вырабатывается защитная реакция;
могут переносить вирусы, бактерии, спирохеты, риккетсии, простейших
Вирусные заболевания.
Спонтанная зараженность иксодовых клей – более 20%. Клещевой весенне-летний энцефалит (Павловский и ученики). Впервые публикация была в 1932 году. В странах Европы – позднее.
Эндемичные очаги приурочены к таежным и лесным зонам, основной переносчик в лиственных и смешанных лесах – собачий клещ, в таежных – таежный.
Распространение энцефалита привело к появлению его в лесостепной зоне.
Клещи – переносчики и природные резервуары. Вирус поражает различные клетки, в т.ч. слюнные половые железы клеща. Через покровы тела передается вирус клещевого энцефалита. Возможна трансовариальная передача вируса. Люди заражаются при укусе или при употреблении сырого зараженного молока. В природных очагах циркуляция вируса осуществляется между клещами и дикими животными – более 22 видов млекопитающих, более 15 видов птиц. Клещевой энцефалит поражает ЦНС человека. У больных наблюдается тошнота. Рвота, сильная головная боль, особенно при поворотах головы, нарушение подвижности языка, потом – потеря сознания, бред, сонливость. Без лечения летальные исходы составляют 30%
Бактериальные заболевания.
Туляремия. Переносчики – разные кровососущие насекомые. Специфические – иксодовые и аргазовые клещи. Напоминает легкие не летальные формы чумы. Может протекать в бубонной форме. Когда бактерии попадают в кровь, там происходит их размножение, что приводит к интоксикации, нарушению функции сердечно-сосудистой системы. Может приобрести хронический характер. Может повторяться через 15-20 лет.
Спирохетозы. Сильно видоизмененные бактерии – спирохеты. Наибольшее значение имеет клещевой возвратный тиф. Передается при укусе и трансовариально. Поселковый клещ – переносчик 20 видов спирохет. Заражение клещей происходит при сосании крови у больных животных. Поражаются легкие, ЦНС, появляются галлюцинации, бред.
Риккетсиозы. Бактериальные микроорганизмы, облигатные внутриклеточные паразиты членистоногих. Перенос осу3ществляется при кровососании. Поражаются кровеносные сосуды, нервная система, отмечаются лихорадки, бред.
169.Класс Насекомые. Классификация. Характерные черты организации. Отряды, имеющие эпидемиологическое значение. Насекомые-возбудители миазов.
Насекомые произошли от 1 из групп многоножек перешедших к жизни на земле. Организация усложнялась, появлялись приспособления к наземному образу жизни. Насекомых очень много и они распространены повсюду.
Есть насекомые, полезные для человека. Есть временные и постоянные паразиты. Отдел зоологии, изучающий насекомых – этанология.
Тело состоит из 3 отделов : голова, грудь, брюшко. Размеры – 0,1 300мм. Имеются глаза (простые или сложные), разнообразные ротовые аппараты.
Грудь состоит из 3 частей (передняя, средняя, задняя). 3 пары конечностей соответственно отделам груди, могут быть крылья.
Брюшко имеет 12 сегментов. Без конечностей. Дышат при помощи трахей, для облегчения процесса служат дыхальца, располагающиеся на груди и брюшке.
Варианты цикла развития:
Яйцо – личинка – куколка – имаго (полный)
Яйцо – личинка – имаго (неполный)
Это самый многочисленный по числу видов класс животных. Общее их количество достигает 1 млн. Тело подразделяют на голову, грудь и брюшко. На голове находятся органы чувств — усики и глаза, сложный ротовой аппарат, строение которого связано со способом питания:
грызущий, лижущий, сосущий, колюще-сосущий и т. п. Грудь насекомых состоит из трех сегментов, каждый из которых несет по паре ходильных ног, построенных по-разному, в зависимости от способа передвижения и двигательной активности. Большинство свободноживу-щих насекомых имеют на груди также две пары крыльев, однако некоторые группы, перешедшие к паразитическому образу жизни, их утратили. Брюшко конечностей не имеет. Органы дыхания насекомых.—трахеи (рис. 21.7). Остальные системы органов насекомых соответствуют организации членистоногих. Развитие насекомых происходит с метаморфозом — неполным,
когда из яйца вылупляется личинка, превращающаяся во взрослую форму или имаго постепенно, после нескольких линек, и полным, при котором в ходе онтогенеза сменяются стадии яйца, личинки, куколки и имаго.
Среди насекомых, имеющих медицинское значение, выделяют следующие группы:
а) синантропные виды, не являющиеся паразитами;
б) временные кровососущие эктопаразиты;
в) постоянные кровососущие паразиты;
г) тканевые и полостные ларвальные (личиночные) паразиты
170.Комнатная муха, муха це-це, вольфартова муха. Систематическое положение, морфология, географическое распространение, развитие, эпидемиологическое значение, меры борьбы и профилактики.
Двукрылые, не включаемые в комплекс гнуса – мухи. Комнатная муха связана с жилищем человека, переносит брюшной тиф, холеру. Дизентерию, туберкулез, полиомиелит, дифтерию, инфекционный конъюнктивит, яйца гельминтов, цисты простейших. На поверхности тела – 60 млн. микроорганизмов, в кишечнике – до 280 млн.
Питается жидкой пищей. Отрыгивает секрет и твердая пища становится жидкой.
Развитие происходит с полным превращением. Яйца откладываются в гниющие остатки. Численность мух обратно пропорциональна культуре человека.
В конце лета – начале осени появляется муха – жигалка. Кровососущая. Просверливает кожу. Нападает на человека и крупных животных. Переносит возбудителей туляремии, сибирской язвы, чумы, бруцеллеза.
Осенняя жигалка Stomoxys calcitrans — муха средних размеров серого цвета со сверлящим хоботком (рис. 21.15, В). Кровью питаются как самцы, так и самки. Личинки развиваются в навозе. В умеренных широтах массовая активность мух наблюдается в конце лета и начале осени, а в тропиках — круглый год. Держится обычно около животноводческих комплексов, активно нападает и на животных, и на человека. Является механическим переносчиком возбудителей туляремии и сибирской язвы.
Для борьбы с массовым выплодом жигалок необходимо гигиеническое содержание домашних животных, а предохранение от укусов достигается индивидуальными средствами защиты.
Муха цеце – специфический переносчик трипаносом.
Некоторым видам мух свойственен – энтомоз – поражение органов и тканей личинками мух (Вольфартова муха).
Мухи це-це р. Glossina широко распространены в экваториальной Африке. Довольно крупные мухи длиной до 13,5 мм. Ротовой аппарат сходен с хоботком осенней жигалки (рис. 21.15, В). Самки живородящи, рождают периодически по одной личинке, которая сразу окукливается, углубляясь в почву. Через 3 недели появляется имагинальная форма. Размножение происходит в тени деревьев и кустов на берегах водоемов. Несколько близких видов мух це-це отличаются друг от друга особенностями окраски, а главное — биологии: одни виды поселяются преимущественно около жилищ человека и питаются в основном его кровью и кровью домашних животных. Другие — обитают в естественной природе (в саваннах и лесах), предпочитая питаться кровью крупных диких копытных, а человека кусают случайно.Все виды являются специфическими переносчиками возбудителя африканского трипаносомоза (см. разд. 19.3.2). Основной мерой борьбы является индивидуальная защита от укусов.
§
Вши, блохи
Отряд ANOPLURA (вши).
Постоянные эктопаразиты млекопитающих, в том числе и человека. Около 500 видов. На человеке могут обитать 3.
– Pediculus capitis
– Pediculus humanus
– Рhthirus pubis
Тело уплощено в спинно-брюшном направлении. Короткие, но мощные ноги, на последнем членике лапки – коготок, необходимый для прикрепления. Размеры имаго 0,5-6,5 мм. Колюще-сосущий ротовой аппарат. Крылья отсутствуют. Брюшко овальной формы имеет 9 сегментов. Развитие протекает с неполным превращением. Яйца – гниды. Из яиц выходит личинка, отличающаяся от имаго меньшими размерами, отсутствием полового аппарата. Живет 16 дней. Откладывает около 300 яиц. Гниды устойчивы (выживают при температуре -3, 65°). При температуре около 100° – быстро гибнут. При погружении в бензин, керосин до 10 минут.
Лобковая вошь (плащица) вызывает фтириаз. Имеет широкое плоское тело, хорошо развиты конечности. Колюще-сосущий ротовой аппарат. Сидит на хозяине неподвижно, высасывая кровь. Выделяет токсичную слюну – хозяин испытывает нестерпимый зуд. Паразитирует в области ресниц, бровей, лобка. На месте кровососания сохраняются синие пятна (до 15 – 20 дней). Заражение осуществляется через контакты, редко при использовании белья.
Головная и платяная вши. Патологическое стояние – педикулез. Переносчик инфекционных заболеваний. Переносят спирохеты (возбудители брюшного тифа), риккетсии (возбудитель волынской лихорадки и эпидемиологического сыпного тифа). Спирохеты и риккетсии размножаются в организме вшей, заполняют их кишечник. Когда вошь сосет кровь здорового человека, нарушается целостность кожных покровов и фекалии вшей с возбудителями опасных заболеваний попадают в кровь.
Н.Г.Минх и О.О.Мочутковский внесли богатый вклад в изучение возвратного сыпного тифа. Поставили опыт по самозаражению. Эпидемии сыпного тифа вплоть до ½ ХХ века вызывали вымирание населения Европы, это было постоянное заболевание Москвы и Петербурга.
1877 – 1878 – 50000 солдат заболели, 17000 погибли от сыпного тифа.
1910 – 1922 – пандемия сыпного тифа в СССР – 10 млн. человек. В основном заболели мужчины в возрасте 20-30 лет.
К постоянным кровососущим паразитам человека из класса насекомых относятся только вши. Человек для них является единственным хозяином, поэтому и трансмиссивные заболевания, возбудителей которых переносят вши, являются типичными антропонозами.
Вши характеризуются выраженными адаптациями к эктопаразитизму: размеры их невелики, конечности снабжены аппаратом фиксации к коже, волосам и одежде, ротовой аппарат колюще-сосущего типа, цикл развития упрощен (развитие с неполным метаморфозом), все стадии жизненного цикла обитают и питаются на хозяине. Постоянство паразитизма этих организмов сопровождается признаками их общей дегенерации: вши в отличие от большинства насекомых имеют не фасеточные глаза, а простые, конечности не обеспечивают быстрого передвижения, полностью редуцированы крылья.
У человека паразитируют два вида вшей: человеческая Pediculus humanus и лобковая Phthirus pubis. Вид человеческая вошь представлен двумя подвидами: Р.h. Capitis — головная, Р.h. humanus — платяная вошь (рис. 21.16).
Кроме человека головная вошь может поселяться на коже головы африканских человекообразных обезьян — шимпанзе и гориллы, что указывает на древность экологических связей с высшими узконосыми обезьянами. У других животных человеческие вши либо не пьют кровь, либо, будучи голодными, пьют, но потом погибают.
В то же время центрально-американские популяции головных вшей способны переходить от человека к капуцинам — низшим широконосым обезьянам, которых местное население традиционно держит в неволе. У диких капуцинов вши не встречаются. Это свидетельствует о широкой экологической пластичности и эволюционных перспективах данного подвида паразитов.
С возникновением человека современного физического типа и его широким расселением по территории с умеренным и холодным климатом вслед за ним стали распространяться и вши. Ношение одежды из шкур и позже из тканей в новых зонах обитания человека создало новые возможности расселения этих паразитов по телу хозяина. В результате появления адаптации к обитанию на одежде возник новый подвид — вошь платяная, которая отличается от головной рядом морфофизиологических признаков, но свободно скрещивается с ней и дает плодовитое потомство. В соответствии с климатическими особенностями исходного ареала обитания платяная вошь встречается только в странах с холодным и умеренным климатом, а в тропиках лишь в условиях высокогорья. Лобковая вошь встречается реже, чем человеческая, но распространена повсеместно. Обитает на лобке, в подмышечных впадинах, иногда на бровях и ресницах. Паразитирование на человеке головной и платяной вшей называется педикулезом. Зараженность лобковыми вшами носит название фтириаза.
Рис. 21.16. Вши. А — головная; Б — платяная; В — лобковая; Г — гниды вшей
Кроме разных мест обитания на человеке вши отличаются друг от друга морфологическими и физиологическими признаками и особенностями жизненного цикла. Самая крупная вошь — платяная. Ее размеры до 4,7 мм. Головная достигает длины 3 мм, а лобковая — не более 1,5 мм. Платяная и головная вши имеют четко отграниченные друг от друга головку, грудь и брюшко, а у лобковой грудь и брюшко слиты. Платяная вошь живет около 50, головная — около 40, а лобковая — до 30 сут. Головная и платяная вши питаются кровью по 2—3 раза в сутки, а лобковая — почти постоянно малыми порциями. Самка платяной и головной вшей откладывает до 300 яиц за всю жизнь, лобковой — до 50. Яйца приклеиваются к волосам или нитям одежды и называются гнидами. Они очень устойчивы.
Слюна вшей обладает токсическими свойствами. Она вызывает ощущение жжения и зуда. У некоторых людей на укусы этих насекомых могут развиваться аллергические реакции. На месте укусов остаются поверхностные кровоизлияния. Ссадины на местах расчесывания инфицируются и загнаиваются. Волосы на голове при этом склеиваются, и образуется трудноизлечиваемое болезненное состояние — колтун.
Лобковая вошь является только паразитом, а головная и платяная — еще и специфическими переносчиками спирохет Borrelia reccurentis — возбудителей возвратного тифа, риккетсий Ricketsia prowaceki — возбудителей эпидемического сыпного тифа, R. wolhynica — возбудителей волынской лихорадки.
Спирохеты размножаются и развиваются в полости тела вшей, поэтому заражение происходит при раздавливании паразитов и попадании их гемолимфы в ранку от укуса или в расчесы.
Риккетсий размножаются в стенке кишечника вшей и выделяются оттуда с фекалиями. Заражение человека происходит при попадании фекалий вшей с риккетсиями в кожные ранки или на слизистые оболочки глаз и дыхательных путей. В связи с тем что риккетсий сохраняют жизнеспособность в сухом виде в течение нескольких месяцев, возможно заражение человека ими без укуса вшами, например при контакте с одеждой больных, в складках которой сохранились сухие фекалии или мертвые насекомые.
Профилактика описанных инфекционных заболеваний — это в первую очередь борьба с педикулезом. Основная мера профилактики педикулеза — соблюдение правил личной гигиены, особенно в местах массового скопления людей. Из химических средств уничтожения вшей используют мази и шампуни, содержащие инсектициды, а также лекарственные препараты, применяемые внутрь и действующие через кровь, которой питаются эти паразиты. Наиболее эффективны короткая стрижка для уничтожения гнид и обработка белья в дезинфекционных камерах.
ОТРЯД APHANIPTORA
Блохи.
Включает более 1000 видов и подвидов. В России встречаются более 500 видов. Облигатные гематофаги. Обитают только на теплокровных, в основном, на млекопитающих. Являются переносчиками инфекционных заболеваний. Размеры 1-5 мм, развитие происходит с полным превращением. Тело сплющено с боков, покрыто прочной хитиновой кутикулой (черного или коричневого цвета). Крыльев нет, конечности развиты (3 пары – хорошо развитые прыгательные ноги). Откладывают яйца там, где много органических веществ (гнезда, трещины полов и др.). 400-500яиц, за кладку – 6 – 10 яиц. Период яйцекладки связан с питанием кровью – гонадотрофический цикл. Эмбриогенез длится 3 – 15 дней. Не могут существовать долго без хозяина. Единственная пища – кровь. В отсутствие крови в коконе не куколка, а блоха. Стимул для выхода из кокона – малейшие сотрясения пола, приводящие к выходу блохи и нападению ее на животное. Капли непереваренной крови все время выступают из анального отверстия и засыхают – это пища для личинок. Когда частицы крови сваливаются с волосами, формируется колтун. Блохи подвижны, человеческая блоха способна подпрыгнуть на 35 см.
Переносчики бактерий (возбудитель чумы, туляремии), вирусов (энцефалит), спирохет (эндемический возвратный тиф). Заражение происходит при кровососании. При проколе кожных покровов вносит возбудителей вместе со слюной. В природе очаги чумы поддерживаются звеньями: грызуны – блохи – грызуны. Блохи являются специфическими переносчиками.
Зарегистрированы 124 вида блох, из которых был выделен вирус чумы в природных условиях. Начало болезни внезапно, начинается подъем температуры, слабость, признаки чумы проявляются на 2 день. Существуют бубонная форма чумы, легочная форма (100% смертность), кожная (происходят кровоизлияния, вся кожа становится темно-синей, происходит сепсис и общее заражение крови) – черная смерть. Известно около 50 государств, где обнаружены или предполагаются природные очаги чумы. Случаи заболевания есть и теперь. Чаще всего заболевания ограничиваются единичными случаями, эпидемий нет. Внимание к чуме не снижается, это – одна из катастроф человечества.
1 пандемия чумы в 542 году в Египте, Сирии. Умирало 5 – 10 тысяч человек в день на протяжении 4 месяцев!
В 14 веке «черная смерть» проникла в Европу из Азии.
1348г на Кипре полностью вымерло население, в Европе погибла ¼ населения, 25 млн. человек.
1351 год – болезнь проникла в Россию. В Смоленске выжило 10 человек.
1754 год – 100000 человек умерло от чумы в Москве.
Зарегистрированы 15 крупных эпидемий чумы в России.
1896 год – Хавкин разработал противочумную вакцину и испытал ее на себе.
Созданы специальные карантинные службы, мешающие перевозке животных, больных чумой. Созданы специальные службы – СЭС и ГосЭпидНадзор. Работа противочумных станций в настоящее время заброшена.
Это мелкие насекомые длиной от 1 до 5 мм. Паразитирование блох облегчается сплющенностью тела с боков, наличием на поверхности его большого количества щетинок, направленных остриями назад, и колюще-сосущим ротовым аппаратом. Задние конечности удлинены и служат для передвижения прыжками. Признаками дегенерации являются рудиментарные глаза и отсутствие крыльев. Развитие блох идет с полным метаморфозом.
Рис. 21.11. Блохи. А—человеческая; Б—крысиная; В—желудок блохи суслика, блокированный бактериями чумы
Наиболее известны человеческая блоха Pulex imtans и крысиная блоха Xenopsylla cheopis (рис. 21.11, А, Б). Оба вида предпочитают питаться кровью соответственно человека и крыс, но легко переходят также на другие виды животных. Крысиная блоха живет в норах крыс, а человеческая — в трещинах пола, за плинтусами и обоями. Здесь самки откладывают яйца, из которых развиваются червеобразные личинки, питающиеся разлагающимися органическими веществами, в том числе фекалиями взрослых блох. Через 3—4 недели они окукливаются и превращаются в половозрелых насекомых.
Человека блохи посещают ночью. Укусы их болезненны и вызывают сильный зуд. Но основное значение блох в том, что они являются переносчиками бактерий — возбудителей чумы. Бактерии чумы, попав в желудок блохи, размножаются там настолько интенсивно, что полностью закрывают его просвет. Это состояние называют чумным блоком (рис. 21.11, В). Если блоха начинает питаться на здоровом животном или человеке, она, проколов кожу, в первую очередь отрыгивает в ранку бактериальный комочек, благодаря чему в кровь поступает сразу огромное количество возбудителей.
Природным резервуаром чумы являются грызуны — крысы, суслики, сурки и др. Эти животные болеют целым рядом других инфекционных заболеваний: туляремией, крысиным сыпным тифом и т. д. Поэтому блохи известны как переносчики возбудителей и этих природно-очаговых заболеваний. Интересно, что кроме трансмиссивного способа заражения указанными болезнями существуют и другие пути: при контакте с зараженными животными, при питье воды из открытых водоемов и т. п., но при укусе блохой заражение является наиболее вероятным, а клиническая картина — наиболее тяжелой.
Борьба с блохами — содержание жилых помещений и хозяйственных построек в чистоте, применение инсектицидов и различных средств борьбы с грызунами. Дают эффект и меры индивидуальной защиты, например репелленты, которыми пропитывают одежду и постельное белье.
172.Комары. Систематическое положение, морфология, географическое распространение, развитие, медицинское значение, меры борьбы и профилактики.
Отряд Двукрылые. У представителей этого отряда пара крыльев, ротовые аппараты их имеют форму хоботка, предназначенного для высасывания жидкой пищи из растительных или животных тканей либо для слизывания ее с поверхности. Развитие с полным метаморфозом. Наибольшее значение представители этого отряда имеют как временные кровососущие паразиты и переносчики возбудителей трансмиссивных заболеваний. Массовые виды этого отряда, размножаясь в летний период в тундре и тайге в огромном количестве, делают жизнь человека почти невыносимой. Разные виды кровососущих двукрылых объединяются общим понятием гнус. Самцы большинства кровососущих двукрылых питаются соками растений, а самки — преимущественно или исключительно кровью животных и человека. В большинстве случаев откладка яиц самками наступает только после кровососания.
Комары (сем. Culicidae). Имеют тонкое стройное тело, длинные ноги и небольшую головку с ротовым аппаратом в виде длинного хоботка. Распространены повсеместно. Они сохраняют в организме и обладают способностью передавать животным и человеку возбудителей более 50 вирусных, бактериальных и паразитарных заболеваний. После оплодотворения самки активно ищут хозяина-прокормителя и способны обнаруживать его на расстоянии до 3 км с помощью обоняния, а затем и зрения. Больных с высокой температурой комары кусают более охотно.
Слабое локальное освещение в вечерние и предутренние часы стимулирует двигательную активность комаров и поиск ими хозяев. В связи с этим комары в темное время суток часто влетают в освещенные окна и двери жилых и хозяйственных построек. В дневные часы суток при высокой температуре и низкой влажности воздуха комары обычно находятся в укрытиях и не питаются. Указанные особенности поведения комаров отражаются на адаптациях к паразитизму возбудителей заболеваний, передающихся комарами: при большинстве заболеваний этой группы у больных повышается температура в вечерние и ночные часы суток, в это же время в крови обнаруживается максимальная концентрация инвазионных для комаров стадий возбудителей.
Комары откладывают яйца в воду или на влажную почву около воды. Личинки и куколки ведут водный образ жизни, а дышат атмосферным воздухом с помощью трахей. Личинки питаются взвешенными в воде мельчайшими органическими частичками. Наиболее известны комары из родов Culex и Aedes (немалярийные комары) — переносчики возбудителей японского энцефалита, сибирской язвы, желтой лихорадки, а также Anopheles (малярийные комары) — специфические переносчики малярийного плазмодия. Доказано, что восприимчивость комаров к заражению возбудителями малярии определяется генотипически и наследуется моногенно. Малярийные и немалярийные комары легко отличаются друг от друга на всех стадиях их жизненного цикла (рис. 21.13).
Яйца малярийных комаров р. Anopheles располагаются на поверхности воды поодиночке, и каждое снабжено двумя воздушными поплавками. Личинки их плавают в горизонтальном положении под поверхностью воды, а на предпоследнем членике имеют пару дыхательных отверстий. Куколки по форме напоминают запятые, находятся, как и личинки, под водной поверхностью и дышат кислородом воздуха через дыхательные рожки, имеющие форму широких воронок. Взрослые малярийные комары, сидя на предметах, располагаются под углом к их поверхности головкой книзу. Находящиеся по обе стороны от хоботка нижнечелюстные щупики равны ему по длине либо немного короче.
Рис. 21.13. Комары на разных стадиях жизненного цикла.
А — Anopheles sp.; Б— Culex sp.:
I — яйца, II — личинки. III — куколки, IV — имаго; 1—воздушные попалвки, 2—плотикиз яиц,3—дыхательные отверстия, 4—дыхательный сифон, 5—воронкообразные дыхательныерожки,6—цилиндрические дыхательные рожки, 7 — нижнечелюстные щупики
Немалярийные комары pp.Culex и Aedes откладывают яйца, слипающиеся группами в небольшие плотики серо-стального цвета. Личинки располагаются под поверхностью воды под углом к ней и имеют на предпоследнем членике длинный дыхательный сифон. Дыхательные рожки куколок имеют форму тонких цилиндрических трубочек, а нижнечелюстные щупики взрослых комаров коротки и достигают не более трети длины хоботка. Тело немалярийные комары держат параллельно поверхности, на которой сидят.
Борьба с комарами наиболее результативна в отношении водных стадий жизненного цикла — личинок и куколок. Применяются мелиоративные методы — засыпка канав и карьеров со стоячей водой. Возможна обработка ядохимикатами отдельных водоемов с большой концентрацией личинок и куколок, а также мест массовых скоплений половозрелых стадий комаров в дневное время суток (сараи, скотные дворы). Наиболее эффективными являются биологические меры борьбы в сочетании с гидромелиоративными, проводящимися в соответствии с государственными антималярийными программами. Так, в Западном Закавказье удалось быстро снизить численность комаров и заболеваемость населения малярией за счет мелиорации и разведения рыб — гамбузий, питающихся преимущественно личинками двукрылых. Для индивидуальной защиты применяют репелленты и механические средства: марлевые пологи, сетки и т. д.
173.Мошки, мокрецы. Систематическое положение, морфология, географическое распространение, развитие, медицинское значение, меры борьбы и профилактики.
Мошки (сем. Simuliidae). Мелкие кровососущие насекомые длиной 2—6 мм. Ротовой аппарат короткий и очень мощный, предназначен для прокалывания кожи и слизывания крови. Крылья прозрачны, без пятен, конечности короткие и толстые (рис. 21.14, Б). Распространены повсеместно, но особую опасность представляют в Африке и тропической Америке, где являются переносчиками возбудителей онхоцеркоза. Интересно, что мошки кусают человека и животных преимущественно в бедра и боковые поверхности туловища. Именно в этих областях скапливаются микрофилярии онхоцерков, что значительно увеличивает вероятность их передачи.
В большом количестве мошки встречаются поблизости от ручьев и речек с быстрым течением, чистой и прохладной водой. На дне таких водоемов самки откладывают яйца, из которых выходят личинки, ведущие прикрепленный и малоподвижный образ жизни. Через 2—3 недели формируется куколка, и еще через 1 неделю из ее оболочки выходят взрослые мошки. Развитие их идет синхронно, и огромное количество мошек на больших территориях появляются одновременно.
Одним из наиболее экологически чистых методов борьбы с мошками является механическая очистка зон особенно быстрого течения рек и ручьев, где на дне и лежащих предметах скапливаются личинки мошек. При этом они отрываются от места прикрепления и уносятся течением. Многие из них при этом гибнут или поедаются различными хищниками. В редких случаях в очагах онхоцеркоза при высокой концентрации личинок мошек на небольших площадях возможно применение инсектицидов.
Мокрецы (сем. Ceratopogonidae). Самые мелкие из кровососущих двукрылых. Их размеры от 1 до 4 мм. Большинство видов имеют пятнистые крылья, длинные многочленистые усики и колюще-сосущий хоботок (рис. 21.14, В). Обитают повсеместно, но так же, как мошки, в тропиках являются переносчиками возбудителей некоторых филяриатозов.
Размножаются мокрецы в небольших стоячих водоемах или во влажной почве. Развитие от яйца до имагинальной стадии продолжается около 1 мес. На человека нападают преимущественно при высокой влажности воздуха или во время дождя. Против мокрецов наиболее эффективны индивидуальные средства защиты.
174.Москиты. Систематическое положение, морфология, географическое распространение, развитие, медицинское значение, меры борьбы и профилактики.
Москиты (сем. Phlebotomidae). Более мелкие насекомые длиной 1,5—3,5 мм, имеющие короткий хоботок, сильно выступающий в виде горбика грудной отдел тела и обильное опущение тела и крыльев мелкими щетинками (рис. 21.14, А).
Москиты встречаются в тропических и субтропических зонах на всех континентах. Они известны как переносчики разных видов лейшманий, вирусов лихорадки паппатачи и возбудителей ряда других трансмиссивных заболеваний. Вирус лихорадки паппатачи передается в поколениях инвазированных москитов трансовариально. Москиты не способны к больших перелетам, поэтому природные очаги заболеваний, вызываемых перечисленными паразитами, обычно имеют небольшие размеры.
Яйца москиты откладывают в норы грызунов и другие затененные места с большим количеством органического вещества и высокой влажностью. Личинки развиваются около 2 мес, а затем окукливаются. Половозрелые стадии появляются через 10—12 сут.
Рис. 21.14. Мелкие кровососущие двукрылые.
А — москит; Б — мошка; В — мокрец
Борьба с москитами должна вестись комплексно и быть направленной на уничтожение природных очагов лейшманиозов и других трансмиссивных заболеваний: это уничтожение грызунов и мест выплода москитов, обработка инсектицидами поверхностей в хозяйственных постройках и жилищах. Эффективны также индивидуальные средства защиты от укусов.
175. Компоненты гнуса.
Компоненты гнуса.
Семейства:
Cullicidae
Simulidae
Ceratopoganidae
Phlebotomidae
Tabanidae
Временные паразиты, кровью питаются только самки, гонотрофическая гармония.
Клетка – элементарная открытая биологическая система. типы клеточной организации. основные положения клеточной теории
Подборка по базе: Вебер Наука как призвание и профессия.docx, экзамен биология 2021 г..doc, ТЕСТ. Наука, псевдонаука, методы.doc, Реферат биология.docx, Реферат Наука как социальный институт.docx, экзамен биология.docx, Реферат Философия как наука и тип мировозрения.docx, тест емтихан биология жауапсыз.docx, тесты биология.docx, реферат биология.docx
Раздел 2
1 . Клетка – элементарная открытая биологическая система. Типы клеточной организации. Основные положения клеточной теории.
клетка – структурная единица всего живого. клетки раститентй и животных организмов сходны по строению.
Клетки представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка, таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль элементарной структурной, функциональной и генетической единицы.
Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия.
Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой.
этапы развит клет теории: 1665г. Роберт Гук – впервые увид под микроскопом насрезе пробки клетки и назвал их «celula». 1674г. – Антуан Ван Левингук – изучает каплю. находит простейших. изучает различные жидкости организма находит в крови – клетки, в сперме – сперматозоиды. (по началу принял их за простейших сожителей).
1825г. – Карл Бэр – открыл яйцеклетку млекопитающего.
1825г. – Я.Пуркинье – увидел жидкое содержимое клетки которое назвал – протоплазма.
1831г. – Роберт Браун – увидел клеточное ядро.
1838(39)г. – Шлейден – изучает растен. общность стоения.
1838г. – Теодор Ван Шванн – создал 2 положен клет теории.
1858г. – Рудольф Вирхов – формирует положение «клетка от клетки» (мож возникнуть из предшеств клетки)
1850г. – Колликер – открыл митохондрий.
1877-81г. Руссов и Горажанкин – обнаружили цитоплазматические мостики и назвали их плазмадесмы.
1898г. –Камило Гольджи – открыл к. гольджи.
(вообще 3 положения: (1) соотносит клетку с живой природой планеты в целом. (оно утверждает, что жизнь, какие бы сложные или простые формы она ни принимала, в её структурном, функциональном и генетическом отношении обеспечивается в конечном итоге только клеткой. (2) указывает что в настоящих условиях единств способом возникновения новых клеток является деление предсуществующих клеток. (3) это полож соотносит клетку с многоклеточными организмами, для которых характерен принцип целостности и системной оргинизации. )
основ полож соврем клет теор: 1) клетка- основная структурная единица живого.
2) клетки всех организмов гомологичны. (сходны по строению. хим составу. обмену веществ и основным закономерностям процессов жизнедеятельности; что свидетельствует о единстве происхождения всех клеток).
3) новые клетки образ путём деления исходной на 2 дочерние. клетка- единица размножения.
4) клетки многоклет организмов сходны по строению, происхождения и выполняемым функциям. объединяются в ткани которые подчиняются единым механизмам регуляции (межклеточные. гуморальные. нервные.)
5) клетки многоклет организмов тиотентны (Тоесть каждая клетка обладает одинаковым набором генов) -> не дифференцированные клетки могут дифференцироваться (тоесть преобразовываться в любой вид клетки)
однако экспрессия генов в различных клетках отлич что приводит к их дифференцировке.
- По продолжительности жизни и по отношению к делению различают три популяции клеток: стабильная, растущая и обновляющаяся.
Стабильная популяция. Клетки этой популяции имеют наибольшую продолжительность жизни, высокодифференцированы и не способны делиться. К такой популяции относятся нервные клетки, клетки миокарда.
Растущая популяция. Клетки этой популяции высокодиф-ференцированные, имеют большую продолжительность жизни. Они составляют основную массу клеток внутренних органов (печень, поджелудочная железа, и т. д.). Эти клетки не утратили способности к делению, они могут перестраивать свой метаболизм, снижать уровень дифференцировки и делиться.
Обновляющаяся популяция. Среди клеток этой популяции есть два типа: высокодифференцированные и недифференцированные (стволовые или камбиальные клетки).Высокодифференцировнные клетки живут недолго (часы, сутки, месяцы), неспособны к делению,
постоянно отмирают. Например, клетки поверхностного слоя эпидермиса, клетки крови, клетки слизистой оболочки кишечника.
Недифференцированные (стволовые) клетки этой популяции постоянно делятся, дифференцируются и замещают погибшие. Так, стволовые клетки эпидермиса находятся в самом нижнем (мальпигиевом) слое; стволовые клетки слизистой оболочки кишечника – в глубоких отделах кишечных крипт, стволовые клетки крови – в красном костном мозге.
Таким образом, ЖЦ клеток стабильной популяции и дифференцированных клеток обновляющейся популяции равен G0, в их жизненном цикле нет митотического цикла.
ЖЦ стволовых клеток обновляющейся популяции состоит из подготовки к делению и деления, т.е. равен МЦ (периодом G0 в данном случае можно пренебречь, т.к. клетки функционально не активны, находятся в состоянии покоя). Такой ЖЦ также имеют клетки злокачественных опухолей, т. к. они не дифференцируются в нормальные клетки, а снова и снова вступают в деление.
ЖЦ клеток растущей популяции состоит из G0 (G1 S G2 M)
2. Строение плазмолеммы: надмембранный, мембранный и субмембранный комплексы. Медицин-ские аспекты функций плазмолеммы.
Плазмолемма( плазматическая мембрана)-слой липидов из двух рядов молекул, гидрофобные участки которых повернуты друг к другу, а гидрофильные-на поверхности слоя. В состав плазмолеммы входят:
•мембранный комплекс-бислой фосфолипидов и встроенных в него белков. Выделяют белки периферические(по внутренней или наружной пов-ти), интегральные( прочно встроенные в мембрану), полупогруженные(когда один конец на поверхности слоя, а другой в гидрофобном слое) и трансмембранные белки( полностью «прошивают» мембрану). Функция этих белков-рецепторная, структурная, ферментативная, антигенная, транспортная.
•надмембранный комплекс-гликопротеиды, углеводные цепи
•субмембранный комплекс-белковые нитевидные структуры( тонкие фибриллы, микрофибриллы, микротрубочки)
Функции мембраны:
•обособление клеток от внешней среды, защита от механического и химического взаимодействия
•восприятие внеклеточных сигналов( рецепторная) и передача их внутрь клетки
•контактное взаимодействие с межклеточным в-вом и другими клетками при образовании органов и тканей
•соединение с элементами цитоскелета-поддержание формы клеток и движение гиалоплазмы внутри клетки
•контролируемый транспорт в-в
Медицинские аспекты функции плазмолеммы:
1-рецепторы гистосовместимости( переливание крови, трансплантация органов)
2-нарушение структуры белков-рецепторов
3-контактное торможение роста клеток( опухолевых клеток)
4-нарушение транспорта некоторых веществ в клетки( развитие атеросклероза и тд)
3. Строение ядра клетки: кариолемма, кариоплазма, хроматин, ядрышки. Понятие об эухроматине и гетерохроматине. Функции ядра.
Строение и функции клеточного ядра
служит хранилищем наследственной информации, содержащейся в хромосомах.
Ядро ограничено ядерной оболочкой, отделяющей его содержимое (кариоплазму) от цитоплазмы.
Оболочка состоит из двух мембран, разделенных промежутком. Обе они пронизаны
многочисленными порами (слияние внешней и внутренней мембраны), благодаря которым
возможен обмен веществами между ядром и цитоплазмой. Полость ядерной оболочки называется перинуклеарным пространством.
В ядре клетки у большинства эукариот находится от 1 до 7 ядрышек. С ними связаны процессы
синтеза РНК и тРНК. В ядре происходит репликация — удвоение молекул ДНК, а также
транскрипция — синтез молекул РНК на матрице ДНК.
Сборка рибосом также происходит в ядре, в ядрышках.
Основные компоненты ядра – хромосомы, образованные из молекулы ДНК и различных белков.
оит из
1)поверхн аппарата ядра (в нем выдлел: 2 мембраны, перинуклеарн пространств, поровые
комплексы, ламину.) 2) кариоплазмы (нуклеоплазмы) 3) хроматина ( в нём эухроматин и гетерохроматин) 4) ядрышка (грануляр и фибриляр компонент.)
Ядро – это структура клетки которая выполняет функцию хранения и передачи инф, а так же регулирует все жизненные процессы клетки. Ядро несёт в себе генетическую (наследственную) инф в виде ДНК. Ядра обычно имеют шаровидную или яйцевидную форму. Я. окружено ядерн оболочкой. Ядерная оболочка пронизана ядерными порами. Через них ядро обменивается веществами с цитоплазмой(внутр средой клетки). Наружная мембрана переходит в эндоплпзматич ретикулум и может быть усеяна рибосомами. Отношение размеров ядра и клетки зависит от функциональной активности клетки. Большинство клеток одноядерные. Двуядерными могут быть кардиомиоциты. Всегда двуядерны инфузории. В них характерен ядерный дуализм.(то есть ядра различ по строению и финкциям). Малое ядро (генеративное) – диплойдное. Оно обеспечивает только половой процесс у инфузорий. Большое (вегетативное) ядро полиплойдное. Оно регулирует все остальные жизненные процессы. Многоядерными бывают клетки некоторых простейших и клетки скелетной мускулатуры.
П.А.Я. или кариотека ) имеет микроскопическую толщину и поэтому виден в световой микроскоп. Поверхностный аппарат ядра включает:
а)ядерную оболочку, или кариолемму;. б)паровые комплексы; в)периферическую плотную пластинку (ППП), или ламину.
(1) Ядерная оболочка (кариолемма). состоит из 2 мембран – наружной и внутренней, разделённых перинукляеарным пространством. Обе мембраны имеют такое же жидкосто-мозаичное строение, как и плазматическая мембрана, и различаются по набору белков. Среди этих белков имеются ферменты, пере-носчики и рецепторы. Наружная ядерная мембрана является продолжением мембран грЭПС и может быть усеяна рибосомами, на которых идёт синтез белка. Со стороны цитоплазмы наружная мембрана окружена сетью промежуточных (ви-ментиновых) фипаментов. Между наружной и внутренней мембранами находится перинуклеарное пространство -полость шириной 15-40 нм, содержимое которого сообщается с полостями каналов ЭПС. По составу перинуклеарное пространство близко к гиалоплазме и может содержать синтезированные рибосомами белки. Главная функция кариолеммы – изоляция гиалоплазмы от кариоплазмы. Специальные белки ядерных мембран, расположенные в области ядерных пор, осуществляют транспортную функцию. Ядерная оболочка пронизана ядерными порами, через которые осуществляется связь кариоплазмы и гиалоплазмы. Для регуляции такой связи в порах находятся (2) поровые комплексы. Они занимают 3-35% поверхности ядерной оболочки. Число ядерных пор с поровыми комплексами является изменчивой величиной и зависит от активности ядра. В области ядерных пор наружная и внутренняя ядерные мембраны сливаются. Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Типичный поровый комплекс представляет собой сложную белковую структуру – содержит более 1000 молекул белка. В центре поры расположена центральная белковая глобула (гранула), от которой по радиусу отходят тонкие фибриллы к периферическим белковым глобулам, образуя диафрагму поры. По периферии ядерной поры находятся две параллельные кольцевые структуры диаметром 80-120 нм (по одному с каждой поверхности кариолеммы), каждое из которых образовано 8 белковыми гранулами (глобулами).
Белковые глобулы перового комплекса подразделяются на центральные и периферические. С помощью периферических глобул осуществляется транспорт макромолекул из ядра в гиалоплазму. (фиксируются в мембране специальным интегральным белком. От этих гранул к центру сходятся белковые фибриллы, формирующие перегородку – диафрагму поры)
В нем участвуют специальные белки периферических глобул – нуклеопорины. В периферических глобулах имеется особый белок – переносчик молекул т-РНК
Центральная глобула специализируется на транспорте и-РНК из ядра в гиалопдазму. В её составе имеются ферменты, участвующее в химической модификации иРНК – ее процессинге.
Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта в-в между цитоплазмой и ядром.
2. Активный перенос в ядро белков
Перенос в цитоплазму субъединиц рибосом Хроматин представляет собой дезоксирибонуклеопротеид (ДНП) и состоит из ДНК, соединённой с белка-ми-гистонами или негистоновыми белками. Гистоны и ДНК объединены в структуры, которые называются нуклеосомами. Хроматин соответствует хромосомам, которые в интерфазном ядре представлены длинными перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Реализацию генетической информации осуществляют деспирализованные участки хромосом.
классификация хроматина:
1) эухроматин (активный деспирализованный. на нем происход считывание инф (транскрипция). в ядре выявляется как более светлые участки ближе к центру ядра) Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна. Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции.
2) гетерохроматин (нерабочий спирализованный, конденсированный, более компактный В ядре выявляется в виде глыбок на периферии.) делится на: конститутивный (всегда неактивен, никогда не переходит в эухроматин) и Факультативный (при определён условиях или на определен стадиях иммунного цикла может переходить в эухроматин). располагается ближе к оболочке ядра, более компактный. Примером скопления факульт гетерохроматина является тельце Барра – инактивированная Х-хромосома у самок млекопитающих, которая в интерфазе плотно скручена и неактивна.
Таким образом, по морфологическим признакам ядра (по соотношению содержания эу- и гетерохроматина) можно оценить активность процессов транскрипции, а, следовательно, синтетической функции клетки.
Хроматин и хромосомы представляют собой дезоксирибонуклеопротеиды (ДНП), но хроматин – это рас-крученное, а хромосомы – скрученное состояние. Хромосом в интерфазном ядре нет, хромосомы появляются при разрушении ядерной оболочки (во время деления)
4.Классификация органоидов клетки по строению и по функции. Строение и функции мембранных органоидов клетки: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, пероксисомы.
Органеллы (organellae) являются обязательными микроструктурами для всех клеток, выполняющими определенные жизненно важные функции. Различают мембранные и немембранные органеллы.
К мембранным органеллам, отграниченным от окружающей их гиалоплазмы мембранами, относятся эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии.
Эндоплазматическая сеть представляет собой единую непрерывную структуру, образованную системой цистерн, трубочек и уплощенных мешочков. На электронных микрофотографиях различают зернистую (шероховатую, гранулярную) и незернистую (гладкую, агранулярную) эндоплазматическую сеть. Внешняя сторона зернистой сети покрыта рибосомами, незернистая лишена рибосом. Зернистая эндо-плазматическая сеть синтезирует (на рибосомах) и транспортирует белки. Незернистая сеть синтезирует липиды и углеводы и участвует в их обмене (например, стероидные гормоны в корковом веществе надпочечников и клетках Лейдига (сустеноцитах) яичек; гликоген – в клетках печени). Одной из важнейших функций эндоплазматической сети является синтез мембранных белков и липидов для всех клеточных органелл.
комплекс Гольджи представляет собой совокупность мешочков, пузырьков, цистерн, трубочек, пластинок, ограниченных биологической мембраной. Элементы комплекса Гольджи соединены между собой узкими каналами. В структурах комплекса Гольджи происходят синтез и накопление полисахаридов, белково-углеводных комплексов, которые выводятся из клеток. Так образуются секреторные гранулы. Комплекс Гольджи имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток комплекс Гольджи расположен вокруг или вблизи ядра, в экзокринных клетках – над
ядром, в апикальной части клетки. Внутренняя выпуклая поверхность структур комплекса Гольджи обращена в сторону эндоплазматической сети, а внешняя, вогнутая, – к цитоплазме.
Мембраны комплекса Гольджи образованы зернистой эндоплазматической сетью и переносятся транспортными пузырьками. От внешней стороны комплекса Гольджи постоянно отпочковываются секреторные пузырьки, а мембраны его цистерн постоянно обновляются. Секреторные пузырьки поставляют мембранный материал для клеточной мембраны и гликокаликса. Таким образом обеспечивается обновление плазматической мембраны.
Лизосомы представляют собой пузырьки диаметром 0,2-0,5 мкм, содержащие около 50 видов различных гидролитических ферментов (протеазы, липазы, фосфолипазы, нуклеазы, гликозидазы, фосфатазы). Лизосомальные ферменты синтезируются на рибосомах зернистой эндоплазматической сети, откуда переносятся транспортными пузырьками в комплекс Гольджи. От пузырьков комплекса Гольджи отпочковываются первичные лизосомы. В лизосомах поддерживается кислая среда, ее рН колеблется от 3,5 до 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Нарушение проницаемости лизосомальной мембраны приводит к активации ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Во вторичных (зрелых) лизосомах (фаголизосомах) происходит переваривание биополимеров до мономеров. Последние транспортируются через лизосомальную мембрану в гиалоплазму клетки. Непереваренные вещества остаются в лизосоме, в результате чего лизосома превращается в так называемое остаточное тельце высокой электронной плотности.
Митохондрии (mitochondrii), являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в формы, доступные для использования клеткой. Их основные функции – окисление органических веществ и синтез аденозинтрифосфорной кислоты (АТФ). Много крупных митохондрий в кардиомиоцитах, мышечных волокнах диафрагмы. Они расположены группами между миофибриллами, окружены гранулами гликогена и элементами незернистой эндоплазматической сети. Митохондрии являются органеллами с двойными мембранами (толщина каждой около 7 нм). Между наружной и внутренней митохондриальными мембранами расположено меж-мембранное пространство шириной 10-20 нм.
Митохондрии – микроскопические двумембранные полуавтономные органоиды общего назначения, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот. Присутствуют во всех эукариотических клетках. В прокариотических клетках митохондрий нет, их функцию выполняют мезосомы – впячивания наружной цитоплазматической мембраны внутрь клетки.
Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Количество митохондрий в клетках, выполняющих различные функции, варьирует в широких пределах – от 50 и достигая в наиболее активных клетках 500-5000. Их больше там, где интенсивны синтетические процессы (печень) или велики затраты энергии (мышечные клетки). В клетках печени (гепатоцитах) их число составляет 800. а занимаемый ими объем равен примерно 20% объема цитоплазмы. Размеры митохондрий составляют от 0,2 до 1-2 мкм в диаметре и от 2 до 5-7 (10) мкм в длину. На светооптическом уровне митохондрии выявляются в цитоплазме специальными методами и имеют вид мелких зерен и нитей (что обусловило их название – от греч. mitos – нить и chondros – зерно).
В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл) органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС). Согласно одной из гипотез, все митохондрии клетки связаны друг с другом и образуют трехмерную сеть.
Митохондрия окружена двумя мембранами – наружной и внутренней, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны – кристы.
* Наружная митохондриальная мембрана гладкая, по химическому составу сходна с наружной цитоплазматической мембраной и обладает высокой проницаемостью для молекул массой до 10 килодальтон, проникающих из цитозоля в межмембранное пространство. По своему составу она похожа на плазмалемму, 25% составляют белки, 75% липиды. Среди липидов присутствует холестерол. Наружная мембранаа содержит много молекул специализированных транспортных белков (например, поринов), которые формируют широкие гидрофильные каналы и обеспечивают ее высокую проницаемость, а также небольшое количество ферментных систем. На ней находятся рецепторы, распознающие белки, которые переносятся через обе митохондриальные мембраны в особых точках их контакта – зонах слипания.
* Внутренняя мембрана имеет выросты внутрь – гребни или кристы, делящие матрикс митохондрии на отсеки. Кристы увеличивают площадь поверхности внутренней мембраны. Таким образом, внутреняя митохондриальная мембрана по площади превосходит наружную. Кристы расположены перпендикулярно или продольно длине митохондрии. Кристы по форме могут быть везикулярные, тубулярные или ламеллярные.
Химический состав внутренней мембраны митохондрий сходен с мембранами прокариот (например, в ней присутствует особый липид – кардиодипин и отсутствует холестерол). Во внутренней митохондриальной мембране преобладают белки, составляющие 75%. Во внутреннюю мембрану встроены белки трех типов (а) белки электрон-транспортной цепи (дыхательной цепи) – НАД’Н-дегидрогеназа и ФАД’Н дегидрогеназа – и другие транспортные белки, (б) грибовидные тельца АТФ-синтетазы (головки которых обращены в сторону матрикса) и (в) часть ферментов цикла Кребса (сукцинатдегидрогеназа). Внутренняя митохондриальная мембрана отличается чрезвычайно низкой проницаемостью, транспорт веществ осуществляется через контактные сайты. Низкая проницаемость внутренней мембраны для мелких ионов из-за высокого содержания фосфолипида
Митохондрии – полуавтономные органоиды клетки, т.к. содержат собственную ДНК, полуавтономную систему репликации, транскрипции и собственный белоксинтезируюший аппарат – полуавтономную систему трансляции (рибосомы 70S типа и т-РНК). Благодаря этому митохондрии синтезируют часть собственных белков. Митохондрии могут делиться независимо от деления клетки. Если из клетки удалить все митохондрии, то новые в ней не появятся. Согласно теории эндосимбиоза митохондрии произошли от аэробных прокариотических клеток, которые попали в клетку хозяина, но не переварились, вступили на путь глубокого симбиоза и постепенно, утратив автономность, превратились в митохондрии.
Митохондрии – полуавтономные органоиды, что выражается следующими признаками:
1) наличие собственного генетического материала (нити ДНК), что позволяет осуществлять синтез белка, а также позволяет самостоятельно делиться независимо от клетки;
2) наличие двойной мембраны;
3) пластиды и митохондрии способны синтезировать АТФ (для хлоропластов источник энергии – свет, в митохондриях АТФ образуется в результате окисления органических веществ).
Функции митохондрий:
1) Энергетическая – синтез АТФ (отсюда эти органоиды и получили название «энергетических станций клетки»):
При аэробном дыхание на кристах происходит окислительное фосфорилирование (образование АТФ из АДФ и неорганического фосфата за счет энергии, освободившейся при окислении органических веществ) и перенос электронов по электрон-транспортной цепи. На внутренней мембране митохондрии расположены ферменты, участвующие в клеточном дыхании;
2) участие в биосинтезе многих соединений (в митохондриях синтезируются некоторые аминокислоты, стероиды (стероидогенез), синтезируется часть собственных белков), а также накопление ионов (Са2 ), гликопротеидов, белков, липидов;
3) окисление жирных кислот;
4) генетическая – синтез нуклеиновых кислот (идут процессы репликации и транскрипции). Митохондриальная ДНК обеспечивает цитоплазматическую наследственность. Аппарат Гольджи (комплекс Гольджи) — мембранная структура эукариотической клетки, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Комплекс Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1898 году. Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок.
Аппарат Гольджи асимметричен — цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭР), на мембранах которого и происходит синтез белков рибосомами. Перемещение белков из эндоплазматической сети (ЭПС) в аппарат Гольджи происходит неизбирательно, однако не полностью или неправильно свернутые белки остаются при этом в ЭПС. Возвращение белков из аппарата Гольджи в ЭПС требует наличия специфической сигнальной последовательности (лизин-аспарагин-глутамин-лейцин) и происходит благодаря связыванию этих белков с мембранными рецепторами в цис-Гольджи. В цистернах Аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам органеллы, в которых происходит их модификации — гликозилирование и фосфорилирование.
ФУНКЦИИ: 1) синтез полисахаридов 2) модификация и окончат созрев всех органич в-в. 3) синтез сложных молеку.(гликолипиды и гликопротеиды. 4) секреция с помощью пузырьков Гольджи. (выводят из клетки синтезир в-во). 5) обновление мембран. (когда секретор пузырёк встраивается в плазмолемму его содержимое вывод их клетки, а сам он становится частью мембраны). 6) участие в выделении растительной клетки. 7) сигрегация (разделение синтезированных в-в на 3 основных потока) это: -собств структура клетки (мембр белки); -секреторные белки; – ферменты лизосом. 8) формирование первичных лизосом.
Впервые эндоплазматический ретикулум был обнаружен американским учёным К. Портером в 1945 году посредством электронной микроскопии. Эндоплазматический ретикулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС) — внутриклеточный органоид эукариотической клетки, представляющий собой разветвленную систему соединённых между собой каналов и полостей, ограниченных одинарной мембраной, поверхность которой составляет более 50% площади всех клеточных мембран. Мембрана ЭПС тоньше чем плазмалемма и содержит более высокую концентрации. белка. Непосредственным продолжение ЭПС является наружная ядерная мембрана.
На поверхности мембран ЭПС происходит большая часть реакций метаболизма, протекающих в клетке. ЭПС разделяет цитоплазму на отдельные отсеки. по каналам ЭПС происходит упорядоченный обмен веществами и энергией между различными компонентами клетки.
ЭПС – генератор мембран для плазмолеммы, ап гольджи и лизосом.
Гранулярная или шероховатая эпс.
наружная обращеная к цитоплазме, сторона грЭПС покрыта рибосомами (котор имеют вид мелк гранул; поступают из ядра благодаря связи мембраны с наруж мембр ядра).
грЭПС – образ уплощенными мембранными цистернами и трубочками на наружной поверхности которых располог рибосомы и полисомы, придающие мембране зернист вид.
Мембраны содерж белки (которые обеспеч связывание рибосом, уплощение цистерн).
Полость грЭПС сообщ с перенуклеарн пространство. Благодаря грЭПС происход отделение вновь синтезированных белковых молекул от гиалоплазмы.
грЭПС хорошо развита в клетках, специализирующихся на белковом синтезе.
ФУНКЦИИ: 1)биосинтез всех мембранных белков, предназначенных для экспорта из клетки.
2) в грЭПС происход посттрансляционный процессинг белков. (созревание белка). белки приобрет характер для них третичную или четвертичную структуру. потом транспортир в комплекс гольджи – > потом в другие органойды.
3) гЭПС выполняет ф-ю пространственного разделения ферментных систем. резделени клетки с помощью мембран на отдел отсеки – компарменты.
4) обеспеч транспорт синтезируемых веществ в аппарат гольджи.
Гладкая или агранулярная ЭПС.
не имеет рибосом. Сост из сильно ветвящихся канальцев и мелких вакуолей диаметром 20-100 нм. гЭПС – трёхмерная замкнутая сеть мембранных анастамозирующих турбочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутсвт.
На цитоплазмотической поверхности гЭПС синтезируется большая часть липидов клетки, которые вход в состав всех её мембран. Часть синтезир на гЭПС белков и липидов встраивается в неё, но увеличения общей площади мембраны при этом не происход. на гЭПС соверш синтез и распад многих углеводов, включ полисахариды, образ стеройдные гормоны.
В Гэпс накаплив многие ядовит в-ва, подлежащ удален из клетки.
гЭПС наиболее развита в клетках с интенсивным жировыми углеводным обменом.
ФУНКЦИИ: 1) синтез липидов; (на мембранах) 2) синтез гликогена (в клетках печени)
3) синтез холестерина и других стеройдов 4) детоксикация эндогенных и экзогенных в-в. (в клетках печени) 5) накопление ионов Са. гЭПС в Миш клетках играет роль депо ионов кальция, необходимых для мыш сокращ. 6) компартментализация (эпс раздел клетку на отдел отсеки) 7) транспорт синтезируемых веществ 8) в мегакариоцитах элементы гЭПС образуют демаркационные каналы, разделяющие формирующие тромбоциты. 9) восстановление кариолеммы в телофазе митоза.
Мембранные органоиды:





