

Практическое применение микроорганизмов
ОБЩАЯ МИКРОБИОЛОГИЯ1
Роль микроорганизмов в природе.
Микроорганизмы – это группа настолько маленьких живых организмов, что они не видны человеческим зрением. Их размер меньше 1 миллиметра, а, порою, намного меньше. Хотя встречаются в этой группе и относительно большие организмы, их даже можно рассмотреть при должном усердии. Изучением их всех занимается наука микробиология
Представители микроорганизмов
В природе существуют десятки тысяч видов микроорганизмов, и это только те, о которых мы знаем. Они являются довольно разнообразными. Некоторые различаются средой обитания, другие – образом или условиями существования, третьи – строением. Так, практически все они одноклеточные, но встречаются среди них и многоклеточные, хоть и редко.
Все микроорганизмы можно поделить на 2 группы: безъядерные (прокариоты) и обладающие клеточным ядром (эукариоты).
Прокариоты – это одноклеточные живые организмы, не имеющие клеточного ядра. Иногда их называют доядерными, а в 20-ых веках именовали монерами, но сейчас этот термин не используется. Представителями прокариотов являются всего 2 домена живых организмов: бактерии и археи. Полагают, что существуют миллионы видов микроорганизмов, относящихся к этим доменам. Но найти то, что не видишь невооружённым взглядом, довольно сложно, поэтому на сегодняшний день известно около десятка-другого тысяч видов.
Микроорганизмы, относящиеся к домену Эукариоты, могут быть как многоклеточными, так и одноклеточными. Главная их особенность – наличие ядра в клетке, поэтому их также называют ядерными. К слову, практически все живые организмы, имеющиеся в природе, относятся к Эукариотам. Исключением являются бактерии, археи, и вирусы.
Микроорганизмы, обладающие клеточным ядром, не очень многочисленны. Их объединяются в одну группу – протисты, хотя входят туда не все ядерные микроскопические организмы.
Бактерии: описание и типы
Бактерии – это домен микроорганизмов (чаще всего одноклеточных), не имеющих ядра. В природе их существует огромное количество видов, сотни тысяч, и даже миллионы, вероятно.
Дело в том, что они настолько малы, что их довольно трудно находить, и уж тем более изучать. Описано лишь около десяти тысяч видов бактерий. Ну а их количество, понятное дело, даже не поддаётся исчислению. Но можно сказать, что практически все из них выполняют некую роль в природе, и обладают некими уникальными свойствами. И исходя из этих знаний, делят бактерий на различные типы, классы, семейства и рода.
Археи: описание и типы
Археи представляют собой домен одноклеточных живых организмов, не имеющих ядра и мембранных органелл. Исходя из особенностей видов этих микроорганизмов, делят их на 8 типов (6 основных и 2 предполагаемых). А каждый из типов, в свою очередь, делится на один или несколько классов архей, обладающих уникальными свойствами. Классы включают в себя семейства и роды архей.
Протисты: описание и типы
Протистами называют группу живых организмов, относящихся к домену Эукариоты (их клетки имеют ядро), и не входящих в состав животных, растений, грибов и хромистов. Создана она по “остаточному принципу”. И, соответственно, включает в себя различные живые организмы, сильно отличающиеся друг от друга.
Исходя из их способа перемещения в пространстве, протистов делят на 3 типа. А каждый из этих типов, исходя из иных особенностей видов, делят на классы, семейства и рода:
· – Инфузории
· – Споровики
· – Саркомастигофоры
Роль микроорганизмов в природе
Микроскопические организмы встречаются практически везде, где имеется вода. Оптимальной температурой для них является 0-50 °C (сильно приблизительно), хотя могут они существовать и при более экстремальных температурах. Рекордно высокая температура для них, как и для всех живых существ, составляет 122 °C. Только стоит понимать, что такую температуру выдержит вовсе не каждый представитель данной группы живых существ. Каждый из видов обладает своими особенностями.
Основная роль микроорганизмов в природе заключается в осуществлении обмена веществ. А поскольку обитают они практически повсеместно, то роль эта очень велика. В большинстве случаев они являются редуцентами, то есть, перерабатывают остатки живых существ. Но иногда выполняют роль продуцентов, производя органические вещества из неорганических. А в силу того, что могут эти существа обитать там, где не выживут другие живые организмы, иногда они являются единственными продуцентами экосистем.
Для человека микроорганизмы могут являться как полезными, так и не очень. Например, благодаря им осуществляет самоочищение воды в различных водоёмах. А ещё они принимают участие в круговороте различных элементов: железа, фосфора, серы и других. Это явная польза, если и не прямая, то как минимум косвенная. Но также существует множество видов организмов, приносящих вред. Некоторые, к примеру, загрязняют водоёмы (если вспомнить, что другие их виды занимаются очищением, это можно счесть забавным). А некоторые вызывают порчу продуктов. А есть даже вредители, которые действуют не опосредованно, а прямо. Речь о патогенных микроорганизмах, или условно-патогенных. Они вызывают инфекционные заболевания.
Заключение
Таким образом, микроорганизмы представляют собой невидимые человеческим зрением живые организмы, обитающие практически везде (поскольку вода находится также практически везде), осуществляющие в природе важные функции, и приносящие человеку как вред, так и пользу.
Практическое применение микроорганизмов
Благодаря большому разнообразию синтезируемых ферментов микроорганизмы могут выполнять многие химические процессы более эффективно и экономично, чем если бы эти процессы проводились химическими методами. Изучение биохимической деятельности микроорганизмов позволило подобрать условия для максимальной активности их как продуцентов различных полезных ферментов – возбудителей нужных химических реакций и процессов. Микроорганизмы все шире применяются в различных отраслях химической и пищевой промышленности, сельском хозяйстве, медицине.
В нашей стране создана и успешно развивается новая отрасль промышленности – микробиологическая, все производства которой базируются на деятельности микроорганизмов.
Микроорганизмы, с помощью которых производят пищевые продукты, называют культурными. Их получают из чистых культур, которые выделяют из отдельных клеток. Последние хранят в музейных коллекциях и снабжают ими различные производства.
В результате осуществляемых культурными микроорганизмами химических реакций растительное или животное сырье превращается в пищевые продукты. С помощью микроорганизмов получают многие жизненно важные продукты питания, и хотя изготовление их знакомо человеку с древних времен, роль в нем микроорганизмов открыта сравнительно недавно.
Хлебопекарное производство.
Хлебопечение основано на деятельности дрожжей и молочнокислых бактерий, развивающихся в тесте. Совместное действие этих микроорганизмов приводит к сбраживанию сахаров муки. Дрожжи вызывают спиртовое брожение, молочнокислые бактерии – молочнокислое. Образующиеся при этом молочная и другие кислоты подкисляют тесто, поддерживая оптимальный для жизнедеятельности дрожжей уровень рН. Углекислый газ разрыхляет тесто и ускоряет его созревание.
Применение культурных микроорганизмов в виде прессованных хлебопекарных дрожжей, сушеных или жидких заквасок улучшает вкус и аромат хлеба.
Производство сыра.
Сыроделие основано на деятельности многих видов микроорганизмов: молочнокислые (термофильный стрептококк), пропионовокислые бактерии и др. Под действием молочнокислых бактерий происходит накопление молочной кислоты и сквашивание молока, под действием других полезных микроорганизмов созревает сыр. Участвуют в этом процессе также некоторые плесневые грибы. Сычужный фермент и молочнокислые бактерии производят глубокое расщепление белков, сахара и жира. Различные бактерии вызывают накопление в острых сырах летучих кислот, придающих им специфический аромат.
§
Творог, сметану, масло, ацидофилин, простоквашу приготовляют на чистых Культурах с применением различных заквасок. Молоко предварительно пастеризуют. Для производства творога и сметаны применяют мезофильные молочнокислые бактерии; ряженки, варенца и подобных продуктов – термофильные стрептококки и болгарскую палочку; ацидофилина – кислотовыносливые молочнокислые бактерии; кефира – многокомпонентные закваски, состоящие из дрожжей, молочнокислых и часто уксуснокислых бактерий. Для изготовления кислосливочного масла в пастеризованные сливки вносят закваску молочнокислых бактерий и выдерживают до требуемой кислотности.
Пивоваренное, спиртовое, ликеро-водочное и винодельческое производства.
Вино, пиво, квас, водку и другие напитки приготовляют с применением дрожжей, вызывающих спиртовое брожение сахарсодержащих жидкостей. В результате брожения жидкости (сусла, бражки, сока и т. п.) образуется алкоголь, СО2 и незначительные количества побочных продуктов. Подсобную роль выполняют молочнокислые бактерии: они подкисляют среду и облегчают деятельность дрожжей (например, при производстве кваса). В производстве спирта и пива для осахаривания заторов применяют также ферментные препараты грибного и бактериального происхождения.
Квашение и соление.
Сущность этого способа консервирования состоит в создании условий для преимущественного развития одних микроорганизмов – молочнокислых бактерий и подавления развития других – гнилостных бактерий. Заквашивают капусту, огурцы, помидоры, яблоки, арбузы. Применяют этот способ также при закладывании на длительное хранение корма для скота – заквашивается зеленая масса из трав, растительных остатков и др. Этот процесс носит название силосования кормов.
Получение органических кислот.
Уксусную, молочную и лимонную кислоты производят также с помощью микроорганизмов. Молочную кислоту получают способом брожения из сахарсодержащего сырья – патоки, крахмала, молочной сыворотки и др.
Молочнокислые бактерии выращивают на средах, содержащих до 15 % сахара. Выход молочной кислоты достигает 60-70 % массы содержащегося в заторе сахара.
Промышленное получение уксуса для пищевых целей основано на уксуснокислом брожении. Уксуснокислые бактерии в специальных чанах на буковых стружках окисляют поступающую питательную среду – уксусно-спиртовой раствор – до уксусной кислоты.
Лимонную кислоту раньше получали из плодов цитрусовых. В настоящее время ее также получают путем брожения. Возбудителем брожения является гриб Аспергиллус нигер, основное сырье – черная патока. Брожение происходит в растворе с содержанием 15 % сахара в аэробных условиях при температуре около 30 °С. Лимонная кислота используется в кондитерской промышленности, производстве безалкогольных напитков, сиропов, кулинарии и медицине.
Микроскопические грибы, бактерии, водоросли, простейшие. Основные свойства и систематика микроорганизмов.
Задачи систематики показать степень родства между организмами и эволюционную связь. Принципы классификации изучает – таксономия (от греч. taxis-расположение, homos – закон) Основные таксономические единицы в систематики микроорганизмов следующие:
· Царство в основе тип клеточной организации (прокариоты, эукариоты, вирусы)
· Отдел – бактерии
· Класс
· Порядок
· Семейство
· Род
· Вид
· Основная таксономическая единица в микробиологии вид, род, семейство.
Вид – бактерии это совокупность микроорганизмов одного генотипа, которые в одинаковых условиях имеют одинаковые фенотипические признаки. Например: Salmonella typhi (род, вид). В пределах одного вида отдельные признаки. Отдельные свойства могут варьировать, поэтому внутри вида могут вида могут определятся под виды (морфологические варианты, биоварианты, хемоварианты, фаговары, серовары(отличаются по антигенным свойствам)).
Установление принадлежности к роду и виду называется идентификацией бактерий. Для идентификации необходимо изучить все свойства микроорганизмов, обнаружить эти свойства в мире микроба. Идентификация микроорганизма происходит по набору следующих свойств: тинкториальный, кульруральных, биохимических, факторов патогенности, антигенных свойств и отношение к фагам. Для облегчения идентификации предложены специальные идентификационные ключи для определенных групп микроорганизмов это наборы признаков для микроорганизма – семейства, рода и вида. Идентификации подвергаются исключительно чистые культуры организма.
ЧКМ – микробы одного вида выращенные в лабораторных условиях на искусственных питательных средах.
Штамм – чистая культура выделенная из конкретного источника, либо чистая культура выделенная из одного источника, но в разное время.
Клон – чистая культура микробов полученная из одной бактериальной клетки
Популяция микробов – совокупность особей одного вида длительно существующих на определенной территории и изолированных от других особой того же вида, популяция единица эволюции. Например популяция сальмонеллы тифи северных регионов отличается от обитающих в южных регионов.
Генетическая таксономия
Идентификация бактерий на основании генетического родства. В основе определение генетических структур клетки — ДНК, внехромосомных структур – плазмиды, транспазоны. Доказано что состав основавний ДНК витоспецифичны т.е. определяется процентное содержание ГЦ от общего содержание всех оснований это может иметь значении е для определение вида. Определяется сходство или комплементарность кислот между различными организмами методом гибридизации. Устанавливают гомологию последовательности нуклеиновых кислот. Этим методом определяют родство между микроорганизмами. % сходства одного порядка равен 80%, для семейства 90%, рода 95% для вида почти 100%.
Первый труд в котором были описаны и классифицированы бактерий был составлен Берджи в 1923 г. В нем бактерии разделены на 25 группы. Патогенные всего 20 групп. В определителе бактерии разделены на Gracilicutes – тонкостенные, Firmicutes — толстостенные, Teniricutes – мягкотелые.
Микроскопические (плесневые) грибы – низшие растения, не имеющие хлорофилла. Это обширная группа микроорганизмов, обладающих различными свойствами. Все они используют кислород воздуха, поэтому развиваются на поверхности продукта, давая паутинообразные разрастания. Многие из этих организмов имеют промышленное значение и используются для получения ферментных препаратов, органических кислот, антибиотиков и др. Отдельные виды плесневых грибов вызывают порчу пищевых продуктов и некоторых товаров, а также заболевания растений, человека и животных.
§
Тело гриба называется грибницей (мицелием) и состоит из тонких разветвленных нитей – гиф, которые переплетены в виде войлока. Клетки мицелия имеют оболочку, цитоплазму с различными включениями, ядра и ядрышки. От мицелия поднимаются воздушные гифы. Плесневые грибы часто являются многоклеточными организмами и видны невооруженным глазом. В этом случае гифы их разделены перегородками на отдельные клетки (септированы). Грибы с несептированными гифами являются одноклеточными (рис. 6).
Рис. 6. Мицелии грибов (гифы): а – несептированный; б – септированный.
Плесневые грибы могут расти не только на твердых, но и на жидких средах при достаточной аэрации в питательной среде или на ее поверхности. Растут грибы концами разветвлений.
В природе микроскопические грибы очень распространены и обитают в почве, воде, на растениях, животных. Находят их и на пищевых продуктах при длительном хранении.
Размножение микроскопических грибов.
Характерной особенностью грибов является разнообразие способов размножения, среди которых можно выделить две большие группы: бесполое (вегетативное) и половое (воспроизведение).
Вегетативное размножение осуществляется обрывками мицелия и воздушных гиф или путем спорообразования. В этом случае споры образуются внутри особых вздутий на концах гиф – спорангиях (эндоспоры) или снаружи на особых гифах – конидиеносцах – (экзоспоры, или конидии).
При половом размножении плесневых грибов сливаются две клетки, дифференцированные в половом отношении. Далее процесс размножения протекает различно, и по его характеру грибы относят к тому или иному классу. У одних грибов образуется клетка с толстой оболочкой и двойным набором хромосом – зигота. После периода покоя она прорастает в новый мицелий. У других грибов после слияния двух клеток образуется многоклеточное плодовое тело различной величины и формы. Внутри него развиваются сумки (аски) со спорами. После их созревания сумка разрывается, споры высыпаются из нее и прорастают в новый мицелий. У некоторых грибов базидиомицетов споры образуются не в сумках, а вне клетки на специальных выростах базидиях.
Созревшие и осыпавшиеся базидии также прорастают в новый мицелий (рис. 7).

Рис. 7. Способы полового размножения микроскопических грибов: а – сумка (аск) со спорами; б – базидии со спорами; в – зигота.
БАКТЕРИЙ.
Бактерии – это микроорганизмы, состоящие всего из одной клетки. Характерная особенность бактерий – отсутствие четко выраженного ядра. Именно поэтому их называют «прокариоты», что означает – безъядерные. Разные виды бактерий
Палочковидные бактерии называют «бациллами».
Кокки – шаровидные бактерии
Спириллы – спиралевидные бактерии
От формы бактерии зависит ее подвижность и способность прикрепляться к той или иной поверхности.. Считается, что бактерии – самые древние организмы на Земле. Они живут практически везде – в воде, почве, атмосфере и внутри других организмов. Внешний вид Бактерии имеют очень маленькие размеры, и увидеть их можно только в микроскоп. Форма бактерий довольно разнообразна. Наиболее распространенные формы – в виде палочек, шариков и спиралек. Бациллы – палочковидные бактерии Палочковидные бактерии называют «бациллами». Кокки – шаровидные бактерии Бактерии в виде шариков – это кокки. Спириллы – спиралевидные бактерии Бактерии в виде спиралек – это спириллы. От формы бактерии зависит ее подвижность и способность прикрепляться к той или иной поверхности.
Строение бактерий Бактерии имеют довольно простое строение. У этих организмов выделяют несколько основных структур – нуклеоид, цитоплазму, мембрану и клеточную стенку, кроме этого, у многих бактерий на поверхности имеются жгутики. Нуклеоид – это подобие ядра, в нем содержится генетический материал бактерии. Он состоит всего из одной хромосомы, имеющей вид кольца. Цитоплазма окружает нуклеоид. В цитоплазме расположены важные структуры – рибосомы, необходимые бактерии для синтеза белка. Мембрана, покрывающая цитоплазму снаружи, играет важную роль в жизнедеятельности бактерии. Она отграничивает внутреннее содержимое бактерии от внешней среды и обеспечивает процессы обмена клетки с окружающей средой. Снаружи мембрана окружена клеточной стенкой. Количество жгутиков может быть разным. В зависимости от вида на одной бактерии бывает от одного до тысячи жгутиков, но встречаются бактерии и без них. Жгутики нужны бактериям для передвижения в пространстве.
Питание бактерий Для бактерий характерно два вида питания. Одна часть бактерий – это автотрофы, а другая – гетеротрофы. Автотрофы сами создают питательные вещества путем химических реакций, а гетеротрофы питаются органическими веществами, которые создали другие организмы.
Размножение бактерий Размножаются бактерии делением. Перед процессом деления хромосома, расположенная внутри бактерии, удваивается. Потом клетка делится надвое. В результате получается две одинаковые дочерние клетки, каждая из которых получает копию материнской хромосомы.
Значение бактерий Бактерии играют важнейшую роль в круговороте веществ в природе – они превращают органические остатки в неорганические вещества. Если бы не было бактерий, то вся земля покрылась бы поваленными деревьями, опавшими листьями и погибшими животными. В жизни человека бактерии играют двоякую роль. Одни бактерии приносят большую пользу, а другие наносят существенный вред. Многие бактерии являются болезнетворными и вызывают различные заболевания, например такие, как дифтерия, тиф, чума, туберкулез, холера и другие. Однако есть бактерии, приносящие пользу людям. Так в пищеварительной системе человека живут бактерии, которые способствуют нормальному пищеварению. А молочнокислые бактерии издавна используются людьми для производства молочнокислых продуктов – сыров, йогурта, кефира и т.д. При квашении овощей и производстве уксуса бактерии также играют важную роль.
Водоросли — большая группа древнейших растений. Строение их тела и размеры характеризуются огромным разнообразием. Существуют микроскопических размеров одноклеточные, многоклеточные и колониальные формы (в 1-2мкм) и крупные, с различным строением слоевища, достигающие 30-45м. Рассмотрим общую характеристику водорослей.
Общим свойством всех водорослей является наличие хлорофилла. Кроме хлорофилла водоросли могут содержать и другие пигменты (фикоциан, фикоэритрин, каротин, ксантофилл, фикоксантин). Эти пигменты придают водорослям красную, бурую, желто-зеленую окраску, Питание. Наличие пигментов в клетках водорослей обеспечивает аутотрофный тип питания. Однако многие водоросли обладают способностью в определенных условиях переключаться на гетеротрофное питание (эвгленовые — в темноте) либо сочетают его с фотосинтезом.
Классификация водорослей. Количество видов водорослей превышает 40тыс. Однако классификация их не завершена, так как не все формы достаточно хорошо изучены. У нас в стране принято деление водорослей на 10 отделов: синезеленые, пирофитовые, золотистые, диатомовые, желтозеленые, бурые, красные, эвгленовые, зеленые, харовые. Наибольшим числом видов представлены зеленые (13-20 тыс.) и диатомовые (10 тыс.).
Деление водорослей на отделы совпадает обычно с их окраской, связанной, как правило, с особенностями строения клеток и слоевища.
Строение клеток водоросли. Водоросли — единственная группа организмов, среди которых встречаются прокариоты (синезеленые) и эукариоты (все остальные). В ядрах эукариотических водорослей выявлены все структуры, свойственные ядрам других эукариот: оболочки, ядерный сок, ядрышки, хромосомы.
маскируя основную зеленую.
Строение, состав и свойства остальных клеточных компонентов у водорослей характеризуется большим разнообразием. В процессе эволюции естественный отбор сохранил наиболее перспективные формы, в том числе такой тип клеточной организации, который позволил растениям перейти к наземному образу жизни.
Размножение водорослей бывает вегетативным, бесполым (с помощью спор) и половым. У одного и того же вида в зависимости от условий и времени года способы размножения различны. При этом наблюдается смена ядерных фаз — гаплоидной и диплоидной.
Условия жизни и места обитания водорослей. Благоприятными условиями существования водорослей являются: наличие света, источников углерода и минеральных солей, а основной средой обитания для них служит вода. Большое влияние на жизнь водорослей оказывают температура, соленость воды и др.
Большинство водорослей — обитатели пресных и морских водоемов. Они могут населять толщу воды, свободно плавать в ней, образуя фитопланктон, либо поселяются на дне, прикрепляясь к разнообразным предметам, живым и мертвым организмам, образуя фитобентос. Заселяют водоросли и горячие источники, а также водоемы с повышенной соленостью.
Кроме планктонных и бентосных различают водоросли наземные, почвенные, снега и льда, известкового субстрата.
Многие водоросли способны вступать в симбиотические отношения с другими представителями растительного и животного царств. Наибольший интерес представляет симбиоз водорослей с грибами. В этом симбиозе возникает такое биологическое единство двух организмов, которое приводит к появлению третьего — лишайника, отличающегося от обоих.
В качестве представителей отдела зеленых водорослей рассмотрим из одноклеточных хламидомонаду, плеврококк и хлореллу, из многоклеточных нитчатых — улотрикс и спирогиру.
§
Потребность прокариот в питательных веществах
Мономеры, необходимые для построения основных клеточных компонентов, могут быть синтезированы клеткой или поступать в готовом виде из среды. Чем больше готовых соединений должен получать организм извне, тем ниже уровень его биосинтетических способностей, так как химическая организация всех свободноживущих форм одинакова.
Источники углерода
В конструктивном метаболизме основная роль принадлежит углероду, поскольку все соединения, из которых построены живые организмы, — это соединения углерода. Их известно около миллиона. Прокариоты способны воздействовать на любое известное углеродное соединение, т. е. использовать его в своем метаболизме. В зависимости от источника углерода для конструктивного метаболизма все прокариоты делятся на две группы: автотрофы, к которым принадлежат организмы, способные синтезировать все компоненты клетки из углекислоты, и гетеротрофы, источником углерода для конструктивного метаболизма которых служат органические соединения. Понятия “авто-” и “гетеротрофия” характеризуют, таким образом, тип конструктивного метаболизма. Если автотрофия — довольно четкое и узкое понятие, то гетеротрофия — понятие весьма широкое и объединяет организмы, резко различающиеся своими потребностями в питательных веществах.
Наибольшая степень гетеротрофности присуща прокариотам, относящимся к облигатным внутриклеточным паразитам, т. е. организмам, которые могут жить только внутри других живых клеток. Паразитический образ жизни привел к редукции некоторых метаболических путей у этих прокариот, что и обусловило полную их зависимость от метаболизма клетки хозяина.
Другие паразитические прокариотные организмы удается выращивать на искусственных средах, но состав таких сред необычайно сложен. Они содержат, как правило, белки или продукты их неглубокого гидролиза (пептиды), полный набор витаминов, фрагменты нуклеиновых кислот и т. д. Для приготовления питательных сред такого состава используют мясные гидролизаты, цельную кровь или ее сыворотку. Формы, способные расти при создании подходящих условий вне клетки хозяина, называют факультативными паразитами.
Следующую крупную группу прокариот составляют так называемые сапрофиты — гетеротрофные организмы, которые непосредственно от других организмов не зависят, но нуждаются в готовых органических соединениях. Они используют продукты жизнедеятельности других организмов или разлагающиеся растительные и животные ткани. К сапрофитам относится большая часть бактерий. Степень требовательности к субстрату у сапрофитов весьма различна. В эту группу входят организмы, которые могут расти только на достаточно сложных субстратах (молоко, трупы животных, гниющие растительные остатки), т. е. им нужны в качестве обязательных элементов питания углеводы, органические формы азота в виде набора аминокислот, пептидов, белков, все или часть витаминов, нуклеотиды или готовые компоненты, необходимые для синтеза последних (азотистые основания, пятиуглеродные сахара). Чтобы удовлетворить потребность этих гетеротрофов в элементах питания, их обычно культивируют на средах, содержащих мясные гидролизаты, автолизаты дрожжей, растительные экстракты, молочную сыворотку.
Есть прокариоты, требующие для роста весьма ограниченное число готовых органических соединений в основном из числа витаминов и аминокислот, которые они не в состоянии синтезировать сами, и наконец, гетеротрофы, нуждающиеся только в одном органическом источнике углерода. Им может быть какой-либо сахар, спирт, кислота или другое углеродсодержащее соединение. Описаны бактерии из рода Pseudomonas, способные использовать в качестве единственного источника углерода и энергии любое из 200 различных органических соединений, и бактерии, для которых источником углерода и энергии может служить узкий круг довольно экзотических органических веществ. Например, Bacillus fastidiosus может использовать только мочевую кислоту и продукты ее деградации, а некоторые представители рода Clostridium растут только в среде, содержащей пурины. Использовать другие органические субстраты для роста они не могут. Биосинтетические способности этих организмов развиты в такой степени, что они сами могут синтезировать все необходимые им углеродные соединения.
Особую группу гетеротрофных прокариот, обитающих в водоемах, составляют олиготрофные бактерии, способные расти при низких концентрациях в среде органических веществ. Организмы, предпочитающие высокие концентрации питательных веществ, относят к копиотрофам. Если у типичных копиотрофов оптимальные условия для роста создаются при содержании в среде питательных веществ в количестве примерно 10 г/л, то для олиготрофных организмов — в пределах 1—15мг углерода/л. В средах с более высоким содержанием органических веществ такие бактерии, как правило, расти не могут и погибают.
Азот
Азот является одним из четырех основных элементов, участвующих в построении клетки. В расчете на сухие вещества его содержится приблизительно 10%. Природный азот бывает в окисленной, восстановленной и молекулярной формах. Подавляющее большинство прокариот усваивают азот в восстановленной форме. Это соли аммония, мочевины, органические соединения (аминокислоты или пептиды). Окисленные формы азота, главным образом нитраты, также могут потребляться многими прокариотами. Так как азот в конструктивном клеточном метаболизме используется в форме аммиака, нитраты перед включением в органические соединения должны быть восстановлены.
Восстановление нитратов до аммиака осуществляется посредством последовательного действия двух ферментов — нитрат- и нитритредуктазы.
Давно была обнаружена способность отдельных представителей прокариотного мира использовать молекулярный азот атмосферы. В последнее время показано, что этим свойством обладают многие прокариоты, принадлежащие к разным группам: эу- и архебактерии, аэробы и анаэробы, фототрофы и хемотрофы, свободноживущие и симбиотические формы. Фиксация молекулярного азота также приводит к восстановлению его до аммиака.
§
При наступлении для бактерий неблагоприятных условий они способны образовывать споры. Неблагоприятными условиями могут быть отсутствие питательных веществ в среде, изменение ее кислотности, высокие или низкие температуры, пересыхание среды и другое.
Образование спор бактериями — это в первую очередь способ пережить неблагоприятные условия среды. В отличие от других организмов у бактерий спорообразование почти не используется для размножения.
Споры бактерий сохраняют жизнеспособность в весьма неблагоприятных условиях внешней среды. Они способны переживать крайне высокие и низкие температуры, сохранять жизнеспособность на протяжении очень многих лет. Так известны бактерии, споры которых могут прорастать через 1000 лет. У других бактерий споры выдерживают кипячение. Бывает, что споры способны пережить температуру меньше -200 градусов Цельсия.
В те времена, когда жизнь на Земле только появилась, и на ней существовали преимущественно только бактерии, возможно погодные условия могли быстро меняться, становиться весьма суровыми. Чтобы выжить, бактерии эволюционно выработали в себе способность к спорообразованию. На сегодняшний день бактерии могут жить там, где другие организмы выжить не могут.
В спорах бактерий все жизненные процессы почти прекращены, цитоплазмы мало, и она густая. Спора покрыта толстой оболочкой, защищающей ее от разрушающих факторов внешней среды. Однако спора содержит все необходимое (в том числе ДНК бактерии), чтобы в благоприятных условиях прорасти и образовать полноценную бактериальную клетку.
Большинство бактерий образуют споры, которые называют эндоспорами. В основном их образуют палочковидные бактерии. «Эндо» значит «внутри». То есть у большинства бактерий споры образуются внутри клетки. При образовании спор происходит впячивание клеточной мембраны, и внутри бактерии обособляется область — будущая спора. Туда переходит ДНК. Вокруг этой области образуется толстый слой так называемой коры, которая будет защищать спору. С ее внутренней и внешней стороны присутствует мембрана. С внешней стороны от мембраны есть еще несколько оболочек.
У палочковидных бактерий эндоспоры могут образовываться в разных местах клетки. У одних — в середине, у других — ближе к концу, у третьих — у самого края палочки-клетки.
Зеленым цветом обозначена оболочка споры, пространство внутри – цитоплазма
Существуют виды бактерий, которые образуют не эндоспоры, а экзоспоры, цисты и другие формы покоящихся форм. «Экзо» говорит о том, что спора образуется не внутри клетки бактерии, а как бы снаружи от нее. Образование экзоспор происходит путем образования своеобразных почек у клетки. После чего такие почки покрываются толстой оболочкой, превращаются в споры и отделяются.
С помощью спор бактерии не только переживают неблагоприятные условия, но и расселяются, так как споры очень легкие и легко разносятся ветром и водой.
Розовым цветом обозначены бактерии, серо-зеленым – их споры. Видно, что споры меньше бактерий
Способы размножения бактерий.
Размножение микроорганизмов — бинарное деление одноклеточных микроорганизмов (бактерий, риккетсий, простейших, дрожжей), в результате которого образуются две новые дочерние полноценные особи, наделенные генетической информацией материнской клетки. Дрожжеподобные грибы могут размножаться почкованием, спорами; плесневые грибы и актиномицеты размножаются обычно спорами.
– Бактерии. Размножаются простым поперечным делением. Бактерии являются гаплоидными клетками. В состав бактериальной клетки входит капсула, клеточная стенка, цитоплазматическая мембрана, цитоплазма, где располагаются мезосомы, рибосомы, нуклеоид, и включения. Некоторые бактериальные клетки имеют жгутики и образуют споры. В отличие от животных клеток такие внутренние структуры бактериальной клетки, как мезосомы, рибосомы, нуклеоид, не имеют мембран, отграничивающих их от цитоплазмы. По способу питания бактерий делят на автотрофов и гетеротрофов, по способу дыхания — на аэробов и анаэробов.
Актиномицеты. Размножаются спорами и поперечным делением (отшнуровыванием) гиф. Занимают промежуточное положение между грибами и бактериями. Среди лучистых грибов сеть патогенные виды — возбудители актиномикозов. Многие актиномицеты являются продуцентами антибиотиков. (см. Антибиотики). В «Определителе» Берджи актиномицеты названы стрептомицетами.
Дрожжи. Существует 2 вида размножения дрожжей — вегетативное (бесполое) и половое с образованием спор. У большинства видов дрожжей вегетативное размножение осуществляется почкованием, редко делением (Schizosaccharomyces). Аспорогенные. дрожжи размножаются только почкованием. Половое размножение происходит при неблагоприятных условиях, когда дрожжи перестают почковаться и превращаются в сумки (аски) со спорами — аскоспоры. Половой процесс заключается в копуляции (слиянии) 2 вегетативных клеток путем сближения их и образования копуляционного канала, в котором происходит слияние частей плазмы и ядра клеток, называемое кариогамией, с образованием диплоидной зиготы, представляющей 2 клетки, соединенные копуляционным каналом. Редукционное деление, или мейоз, сопровождаемое уменьшением числа хромосом вдвое, происходит сразу, без полового процесса, и зигота превращается в аск с 4 гаплоидными спорами, поэтому вегетативное поколение таких спор гаплоидно. Споры прорастают без копуляции. Так происходит размножение у дрожжей Zygosaccharomyces (рис. 2Б). У дрожжей Saccharomyces половой процесс происходит при слиянии спор или проросших из них клеток с образованием диплоидной зиготы, которая сразу начинает почковаться, образуя диплоидное потомство (рис. 2А). Мейоз происходит непосредственно перед образованием спор.
Плесневые грибы. У Грибов различают вегетативное, половое и бесполое размножение. Вегетативное размножение может осуществляться при отделении от основной массы мицелия его частей, которые могут развиваться самостоятельно, а так же путем почкования мицелия или отдельных клеток у дрожжевых грибов. Половое размножение состоит в слиянии половых клеток, в результате чего возникает зигота. Бесполое размножение осуществляется при помощи специальных образований, называемых спорами. Споры могут развиваться внутри специальных споровместилищ или на концах особых выростов мицелия – конидиеносцах. Основной способ размножения плесневых грибов – при помощи спор. Плесень размножается невероятно быстро.
В обыкновенной хлебной плесени можно различить маленькие чёрные точки – спорангии, в которых образуются споры. В одном спорангии содержится до 50.000 спор, каждая из которых способна воспроизвести сотни миллионов новых спор всего за несколько дней! А если условия благоприятные, плесень быстро появится на книге, обуви или на упавшем дереве в лесу.
§
Размножение бактерий
Наиболее часто бактерии размножаются путем бинарного деления, когда из одной клетки образуется две, каждая из которых вновь делится. Процессу деления всегда предшествует репликация (удвоение) ДНК. Существует два типа деления – деление перетяжкой (перешнуровывание) и с помощью поперечной перегородки (рисунок А.7) .
Деление перетяжкой (констрикция) сопровождается сужением клетки в месте ее деления, и в этом процессе принимают участие все слои клеточных оболочек. Выпячивание оболочек внутрь клетки все более ее сужает и, наконец, делит на две. Это деление присуще грамотрицательным бактериям. Деление с образованием поперечной перегородки присуще грамположительным бактериям. Однако у некоторых групп бактерий отмечена смена способов деления (тионовые бактерии, микобактерии). У шаровидных бактерий может образовываться несколько поперечных перегородок (тетракокки, сарцины). Почкование убактерий является разновидностью бинарного деления. Этот способ размножения присущ бактериям, имеющим диморфные или полиморфные клеточные циклы. Почкующимся бактериям присуща полярность клеток. Некоторые бактерии размножаются с помощью экзоспор (но не эндоспор!), некоторые – фрагментами гиф (актиномицеты). У некоторых бактерий имеются половые ворсинки, или F-пили.
Период от деления до деления называется клеточным циклом. Различают несколько типов вегетативного клеточного цикла: мономорфный – образуется только один морфологический тип клеток (например, бациллы), диморфный – два морфологических типа клеток, полиморфный – несколько (актиномицеты). При диморфном и полиморфном циклах различают дочерние и материнские клетки.
Бактерии характеризуются высокой скоростью размножения. Например, в благоприятных условиях кишечная палочка делится каждые 20-30 мин, за сутки это дает 272, т.е. 72 поколения. В условиях, исключающих гибель, эта биомасса составит 4720 т. Скорость размножения зависит от факторов внешней среды (температуры, условия питания, влажность, реакция среды и др.) и от видовых особенностей бактерий. Высокая скорость размножения бактерий обеспечивает их сохранение на земле даже в условиях массовой гибели. Сохранившиеся отдельные клетки размножаются и вновь дают поколение.
Размножение актиномицетов
Actinomyces размножаются с помощью субстрационного мицелия прорастающего в субстрате и воздушного мицелия вырастающего с плодоносцев-спорангиофоров. Плодоносцы в зависимости от вида имеют различную форму завитков от закрученной формы, до прямой или волнистой.
Некоторые виды актиномицетов имеют спороносные ветки расположенные в виде мутовок или пучков, часто они моноподиально висят на нитях мицелия.
Образование спор происходит с помощью фрагментации или сегментации.
Фрагментация– это процесс дробления протопласта спороносной ветки на сто и более меленьких комков содержащих базофильное а также ядерное вещество. Комки, превращаясь в споры, располагаются длинной цепочкой в спороносце.
Сегментация – это процесс разделения спороносца на сегменты с палочковидной формой, с помощью поперечных перегородок, они округляются и преобразовываются в споры. Оболочки спор у разных видов имеют гладкую иногда бугристую, зубчатую, шиповидную, волосистую поверхность. Выросты на поверхности оболочек хорошо видны через электронный микроскоп.
В большинстве случаев актиномицеты аэрофилы и мезофилы, но получили распространение и термофилы, многие их виды способны образовывать пигменты разных цветов. Актиномицеты, имея разнообразный набор ферментов способны синтезировать различные вещества и выделять их большими количествами в окружающую среду. Среди этих веществ, обладающих высокой физиологической активностью, встречаются многие витамины, некоторые аминокислоты, токсины, каротиноиды, фитогормоны и другие.
Также стоит упомянуть способность актиномицетитов образовывать разнообразные виды антибиотиков.
Метанобразующие бактерии
Предположение о биологической природе образования метана было высказано еще в XIX в. Однако изучение этого процесса и организмов, его осуществляющих, тормозилось из-за отсутствия чистых культур. Сложность заключается в чрезвычайной чувствительности большинства метанобразующих бактерий к О2. Быстрый прогресс в изучении этой группы архебактерий связан с использованием методов культивирования анаэробов, разработанных Р.Е.Хангейтом (R.E.Hungate). В качестве основных приемов используется удаление О2 из газов, в атмосфере которых осуществляются культивирование и все необходимые для работы операции, а также применение предварительно восстановленных сред.
Метанобразующие бактерии (метаногены) – морфологически разнообразная группа, объединяемая двумя общими для всех ее представителей признаками: облигатным анаэробиозом и способностью образовывать метан. Для создания таксономической структуры метанобразующих бактерий был использован филогенетический подход, основанный на сравнительном анализе нуклеотидных последовательностей 16S рРНК. В соответствии с таким подходом в девятом издании Определителя бактерий Берги группа разделена на три порядка ( Methanobacteriales , Methanococcales , Methanomicrobiales ), коэффициент сходства ( SAB ) для которых составляет 0,2-0,28. Далее порядки разделены на 6 семейств (SAB=0,34-0,36) и 13 родов (Sab=0,46-0,51). Число видов достигает более 40. SAB для них колеблется в пределах 0,55-0,65. О гетерогенности группы можно судить и по нуклеотидному составу ДНК ее представителей (молярное содержание ГЦ-оснований – от 27 до 61%).
В состав группы входят бактерии с разной морфологией: прямые или изогнутые палочки разной длины; клетки неправильной формы, близкие к коккам; извитые формы. У некоторых видов наблюдается тенденция формировать нити или пакеты. Клетки неподвижные или передвигающиеся с помощью перитрихиально или полярно расположенных жгутиков. У представителей рода Methanosarcina в клетках найдены газовые вакуоли. Для некоторых метаногенов характерна развитая система внутриклеточных элементарных мембран, являющихся результатом разрастания и впячивания в цитоплазму ЦПМ и сохраняющих с ней связь. У этой группы архебактерий обнаружены клеточные стенки трех типов: состоящие из псевдомуреина, построенные из белковых глобул и гетерополисахаридной природы. Описан микоплазмоподобный метаноген, выделенный в род Methanoplasma , не имеющий клеточной стенки и фильтрующийся через мембранные фильтры с диаметром пор 0,45 мкм.
20-30% мембранных липидов метаногенов представлены нейтральными и 70-80% – полярными липидами. Последние – это в основном два типа простых эфиров глицерина и терпеноидных спиртов (С20-фитаниловый и С40- бифитаниловый), на основе которых образуются полярные фосфо- и гликолипиды (рис. 14 и рис. 102 ). В зависимости от вида клеточные мембраны могут содержать оба типа эфиров или только один. Основными нейтральными липидами являются С20-, С25- и С30-ациклические изопреноидные углеводороды, насыщенные или содержащие двойные связи. Запасных продуктов в виде поли-бета-оксимасляной кислоты или гликогена в клетках не обнаружено.
Метанобразующие бактерии – строгие анаэробы . Первые исследования чистых культур, выделенных из рубца жвачных животных, показали, что рост их возможен при начальном окислительно-восстановительном потенциале среды ниже -300 мВ. Рост некоторых видов полностью подавляется при содержании в газовой фазе более 0,004% молекулярного кислорода. Однако описаны виды с относительно низкой чувствительностью к О2. В их клетках найдена супероксиддисмутаза . Возможно, в природе такие виды могут сохранять жизнеспособность при кратковременных контактах с О2 и возобновлять рост в анаэробных условиях.
Большинство метанобразующих бактерий имеют температурный оптимум для роста в области 30-40 градусов по С, т.е. являются мезофилами , но есть виды, у которых оптимальная зона сдвинута в сторону более низких (25 градусов по С) или высоких (55-65 градусов по С) температур. Выделен экстремально термофильный организм Methanothermus fervidus , растущий при 55-97 градусов по С (оптимум 80 градусов по С). Все известные представители этой группы – нейтрофилы с оптимальным рН в области 6,5-7,5. Среди метаногенов есть галофилы, требующие в качестве одного из оптимальных условий для роста содержания в среде до 65-70 г/л NaCl.
В качестве источника углерода и энергии для роста метаногены используют узкий круг соединений. Наиболее универсальными источниками углерода и энергии для них является газовая смесь Н2 и СО2. Более 3/4 известных видов утилизируют Н2 СО2. Некоторые метаногены приспособились к облигатному использованию этих соединений. Следующими по распространенности источниками углерода и энергии служат формиат , ацетат , метанол , метиламины и моноокись углерода.
Около половины изученных видов не нуждаются в каких-либо органических соединениях. Для роста многих культур в атмосфере Н2 и СО2 требуется внесение в среду органических веществ, стимулирующих рост или абсолютно для него необходимых. Это могут быть некоторые витамины группы В, ацетат, пируват , сукцинат , отдельные аминокислоты, дрожжевой экстракт или компоненты неизвестного состава, содержащиеся в природных средах обитания. Так, штаммы, выделенные из рубца жвачных животных, нуждаются в добавках рубцовой жидкости. Сложные органические соединения метанобразующие бактерии использовать не могут.
В качестве источника азота метаногены используют аммонийный азот или некоторые аминокислоты. Для ряда видов показана способность к азотфиксации. Источником серы могут служить сульфаты , сульфидили серосодержащие аминокислоты.
§
Основными местами обитания метанообразующих бактерий являются осадки пресных, морских, гиперсоленых водоемов, гидротермы, очистные сооружения, заболоченные почвы. Кроме того, эти бактерии являются важными компонентами пищеварительного тракта растительноядных животных; найдены они в толстом кишечнике человека (таблица 2).
Субстратная специфичность метанообразующих бактерий обусловливает необходимость их взаимодействия с другими анаэробными бактериями – продуцентами необходимых метаногенам субстратов. В этой связи метаногены в природе развиваются в составе сложного общества, которое обеспечивает в анаэробных условиях разложение практически любого органического соединения. Конечной стадией такого разрушения является образование метана, который, насколько известно, не может быть использован в анаэробных реакциях.
В основе взаимодействия микроорганизмов, входящих в сообщество, лежит трофическая цепь, ведущая от сложных биополимеров к простым газообразным продуктам. При этом на первом этапе, благодаря жизнедеятельности “первичных анаэробов”, идет деструкция высоко-энергизованных природных соединений с образованием летучих жирных кислот (формиата, ацетата, пропионата, бутирата), органических кислот (лактата, сукцината), низших спиртов (этанола, метанола), альдегидов, кетонов, водорода и углекислого газа. На втором этапе действуют “вторичные анаэробы”, которые используют перечисленные выше продукты в качестве энергетических субстратов. В результате образуется ацетат, Н2, СО2, формиат. В последнюю очередь происходит образование метаногенами метана, который далее удаляется из системы в анаэробную зону в виде пузырьков газа, где используется метанотрофными бактериями.
Согласно современным представлениям процесс образования метана происходит по двум механизмам. Первый механизм начинается с последовательности процессов, приводящих к центроболитам (НСООН, СН3ОН и др.), которые затем используются метанообразующими бактериями. Этот механизм метаногенеза связан с переносом углеродсодержащего материала в трофической цепи.
Второй механизм обусловлен потреблением метаногенами Н2. Следствием этого процесса является изменение метаболизма первичных анаэробов в сторону увеличения выхода энергии. Тесное взаимодействие между микроорганизмами на основе межвидового переноса водорода получило название синтрофии.
Метанообразующие бактерии представляют определенный практический интерес как продуценты газообразного топлива – метана. В институте биохимии им. Баха разработан способ получения в процессе метанового брожения кормового витамина В12 при сбраживании отходов, богатых углеводами.
Метанообразующие бактерии являются одним из основных компонентов активного ила метантенков – анаэробных очистных сооружений сточных вод. В данном случае бактерии используются для сбраживания углеродсодержащих осадков. Многие продукты анаэробного распада в метантенках могут быть утилизированы. Выделяющийся в процессе метанового брожения метан собирается в газгольдеры и используется на отопление очистной станции, в том числе и на поддержание требуемой температуры в метантенке. Сброженный осадок после подсушивания на иловых площадках или механического обезвоживания может быть использован в качестве удобрения на сельскохозяйственных полях.
10 Дыхание микроорганизмов, сущность процесса. Способы дыхания.
Для своей жизнедеятельности каждая микробная клетка кроме питательных веществ нуждается в энергии. Эту энергию микроорганизмы получают в процессе дыхания. Сущность дыхания у микробов заключается в окислении сложных органических соединений до более простых веществ с выделением тепловой энергии, которая и используется микробами. В большинстве случаев микроорганизмы получают энергию путем окисления углеводов и других органических соединений. За счет полученной энергии происходит синтез сложных органических соединений в самой клетке.
По типу дыхания микроорганизмы делятся на аэробы и анаэробы. Аэробы – микроорганизмы, которые для дыхания и получения необходимой энергии нуждаются в свободном доступе кислорода из воздуха. У этой группы микробов процесс дыхания аэробный. Анаэробы – микроорганизмы, которые получают энергию при дыхании без доступа кислорода воздуха путем расщепления питательных веществ. Различают облигатные (строгие) и факультативные (нестрогие) анаэробы. Облигатные анаэробы проявляют свою жизнедеятельность только при отсутствии кислорода воздуха. Факультативные анаэробы могут развиваться в средах как в присутствии кислорода воздуха, так и без него.
В химизме дыхательных процессов у аэробов и анаэробов имеется много общего. Во всех случаях первым этапом дыхательных процессов является отщепление водорода от субстрата (дегидрирование) в присутствии специфических ферментов – дегидрогеназ. Происходящие процессы носят окислительно-восстановительный характер.
Сущность окисления состоит в потере электронов окисляющимся веществом, тогда как сущность восстановления состоит в присоединении этих электронов восстанавливающимся веществом. Та или иная последовательность биохимических реакций в течение обменных процессов возможна благодаря тонким изменениям окислительно-восстановительного потенциала, под которым понимают способность вещества отдавать или получать электроны.
При аэробном типе дыхания аэробные дегидрогеназы передают отнятый от субстрата водород или непосредственно кислороду воздуха, или цитохромной системе. Это так называемое аэробное дегидрирование, при котором происходит обычно полное окисление. При полном окислении конечными продуктами являются вода и углекислота, при этом освобождается вся энергия. При неполном окислении происходит образование продуктов, в которых заключается значительная часть энергии.
В анаэробных условиях биохимические процессы происходят при отсутствии кислорода воздуха. Анаэробные дегидрогеназы не могут отдавать водород кислороду воздуха, а передают его другим веществам, от которых сравнительно легко отщепляется кислород. Это так называемое анаэробное дегидрирование, при котором происходит неполное окисление субстрата.
Дыхание микроорганизмов представляет собой биологическое окисление различных органических соединœений и некоторых минœеральных веществ. В итоге окислительно-восстановительных процессов и брожения образуется тепловая энергия, часть которой используется микробной клеткой, а остальное количество выделяется в окружающую среду. Сегодня окисление определяют как процесс отнятия водорода (дегидрирование), а восстановление — его присоединœения. Эти термины применяют к реакциям, связанным с переносом протонов и электронов или только электронов. При окислении вещества происходит потеря электронов, а при восстановлении — их присоединœение. Считают, что перенос водорода и перенос электронов — эквивалентные процессы.
Энергия, освобождаемая в процессе окислительно-восстановительных реакций, накапливается в макроэргических соединœениях АДФ и АТФ (аденозиндифосфат и аденозинтрифосфат). Эти соединœенияимеют макроэргические связи, обладающие большим запасом биологически доступной энергии. Οʜᴎ локализованы в сложно устроенных структурах микробных клеток — мезосомах, или митохондриях.
По типу дыхания микроорганизмы делят на аэробов, анаэробов и факультативных анаэробов.
Аэробное дыханиемикроорганизмов — это процесс, при котором последним акцептором водорода (протонов и электронов) является молекулярный кислород. В результате окисления главным образом сложных органических соединœений образуется энергия, которая выделяется в среду или накапливается в макроэргических фосфатных связях АТФ. Различают полное и неполное окисление.
Полное окисление.Основной источник энергии у микроорганизмов —углеводы. В результате расщепления глюкозы в аэробных условиях процесс окисления идет до образования диоксида углерода и воды с выделœением большого количества свободной энергии:
С6Н12О6 6О2 → 6СО2 6Н2О 674 ккал.
Неполное окисление.Не всœе аэробы доводят реакции окисления до конца. При избытке углеводов в среде образуются продукты неполного окисления, в которых заключена энергия. Конечными продуктами неполного аэробного окисления сахара бывают органические кислоты: лимонная, яблочная, щавелœевая, янтарная и другие, которые образуются плесневыми грибами. Так же осуществляется аэробное дыхание уксуснокислыми бактериями, в которых при окислении этилового спирта образуется не диоксид углерода и вода, а уксусная кислота и вода:
С2Н5ОН О2 → СН3СООН Н2О 116 ккал.
этиловый спирт уксус. к-та
Окисление этилового спирта уксуснокислыми бактериями может идти и дальше — до появления диоксида углерода и воды, при этом освобождается большое количество энергии:
С2Н5ОН 3О2 → 2СО2 3Н2О 326 ккал.
этиловый спирт
Анаэробное дыханиеосуществляется без участия молекулярного кислорода. Различают собственно анаэробное дыхание (нитратное, сульфатное) и брожение. При анаэробном дыхании акцептором водорода являются окисленные неорганические соединœения, которые легко отдают кислород и превращаются в более восстановленные формы. Нитратное дыхание — восстановление нитратов до молекулярного азота. Сульфатное дыхание — восстановление сульфатов до сероводорода.
Брожение— расщепление органических углеродсодержащих соединœений в анаэробных условиях. Оно характеризуется тем, что последнимакцептором водорода служит молекула органического вещества с ненасыщенными связями. Вещество при этом разлагаетсятолько до промежуточных продуктов, представляющих собой сложныеорганические соединœения (спирты, органические кислоты). Заключенная в них энергия не используется микробами, а об  разовавшаяся в небольших количествах энергия выделяется в окружающую среду.
разовавшаяся в небольших количествах энергия выделяется в окружающую среду.
Типичными примерами анаэробного дыхания являются:
– спиртовое брожение (дыхание дрожжей в анаэробных условиях):
С6Н12О6 → 2С2Н5ОН 2СО2 27 ккал;
этилов. спирт
– молочнокислое брожение (дыхание молочнокислых бактерий):
С6Н12О6 → 2С3Н6О3 18 ккал;
молочн. к-та
– маслянокислое брожение (дыхание маслянокислых бактерий):
С6Н12О6 → С3Н7СООН 2СО2 2Н2 15 ккал;
маслян. к-та
Как видно из приведенных уравнений, при анаэробном дыхании освобождается значительно меньше энергии, чем при аэробном. По этой причине при анаэробном дыхании для того, чтобы обеспечить потребность в крайне важно м количестве энергии, микроорганизмам крайне важно потреблять больше сахаров, чем при аэробном.
Большая часть энергии, образующейся при дыхании, освобождается в окружающую среду. Это вызывает нагревание продуктов, в которых развиваются микроорганизмы. Именно так нагревается вино, в котором происходит спиртовое брожение; нагревается влажное зерно, торф, сено.
§
Разложение целлюлозы осуществляют различные представители бактерий, грибов и простейших. Все эти микроорганизмы называются целлюлозо- литическими. К целлюлозолитическим грибам относятся представители родов Fusarium, Chaetomium и Neocallimastix, а также виды Trichoderma viride, Aspergillus fumigatus, Botrytis cinerea, Rhizoctonia solani, Myrothecium verrucaria. Разложение целлюлозы осуществляют простейшие родов Diplodinium, Eudiplodinium, Entodinium. К целлюлозолитическим бактериям относятся: аэробные, анаэробные виды. Аэробное расщепление целлюлозы осуществляют представители родов Cytophaga, Sporocytophaga, Polyangium, Sporangium, Bacillus, Actinomyces, Alcaligenes, Cellulomonas, Pectobacterium, Erwinia, Microbispora, Pseudomonas, Streptomyces и Thermomonospora. В анаэробных условиях целлюлозу расщепляют представители родов Bacteroides, Butyrivibrio, Clostridium, Fibrobacter, Thermoanaerobacter и Ruminococcus. Разложение целлюлозы с помощью микроорганизмов проходит в несколько этапов. Ферментная система, осуществляющая разложение целлюлозы до глюкозы, носит название целлюлазного комплекса. В его состав входят по меньшей мере три фермента: эндо-β- 1,4-глюканаза, экзо-β-1,4-глюканаза, β-глюкозидаза. Процесс расщепления целлюлозы протекает следующим образом: под действием фермента эндо-β-1,4-глюканазы происходит разрыв различных β-1,4-гликозидных связей внутри макромолекулы с образованием больших фрагментов со свободными концами. Затем под влиянием экзо-β-1,4-глюканазы от концов образованных фрагментов отщепляются дисахариды целлобиоза. β-глюкозидаза осуществляет гидролиз целлобиозы с образованием 2 молекул глюкозы. При аэробном разложении целлюлозы глюкоза далее окисляется в основном до СО2 и Н2О. Могут также накапливаться в небольших количествах органические кислоты. При анаэробном распаде целлюлозы первоначальный продукт ее гидролиза – глюкоза подвергается сбраживанию, в результате чего образуются различные органические кислоты (уксусная, молочная, муравьиная, масляная, янтарная), этанол, СО2, Н2. Состав продуктов брожения отличается у разных видов микроорганизмов. Примером анаэробного сбраживания целлюлозы являются микробиологические процессы, которые осуществляются в рубце жвачных животных.
Пектиновые вещества входят в состав срединных пластинок, образующихся между стенками соседних растительных клеток. Существуют три типа пектиновых веществ: протопектин – водонерастворимая составная часть клеточной стенки; пектин – водорастворимый полимер галактуроновой кислоты, содержащий метилэфирные связи; пектиновая кислота – водорастворимый полимер галактуроновой кислоты, свободный от метилэфирных связей. Микроорганизмы синтезируют следующие ферменты, катализирующие распад пектиновых веществ: протопектиназу, осуществляющую разложение протопектина с образованием растворимого пектина; пектинэстеразу, гидролизующую метилэфирную связь пектина с образованием пектиновой кислоты и метилового спирта; пектиназу (полигалактуроназу), разрушающую связи между отдельными составляющими галактуроновой кислоты, пектина или пектиновой кислоты с образованием свободной D- галактуроновой кислоты. Способность расщеплять пектиновые вещества присуща многим грибам и бактериям. Из бактерий высокой пектолитической активностью обладают представители рода Bacillus (B. macerans, B. polymyxa), рода Clostridium ( С. pectinovorum, C. felsineum, C. pectinolyticum, C. flavum, C. corallinum и др.), рода Pectobacterium (P. carotovorum, P. atrosepticum и др.). Пектиновые вещества разлагаются под влиянием фитопатогенных грибов Botrytis cinerea и Fusarium oxysporum. Следует отметить, что численность пектолитических микро – организмов в почве чрезвычайно велика (10 5 клеток на 1 г почвы) Микроорганизмы, разлагающие пектиновые вещества, играют важную роль при мочке льна, конопли, джута, канатника, кенафа и других лубоволокнистых растений. Цель этого процесса – отделение пучков целлюлозных волокон от остальных растительных тканей, которые склеены пектиновыми веществами. Пектолитические ферменты, кроме того, используются для различных технических целей (например, для осветления фруктовых и овощных соков).
Липиды встречаются в растительных и животных клетках в виде запасов питательных веществ. По химическому составу это чаще всего глицериды – сложные эфиры глицерина и жирных кислот. В почву и водоемы липиды попадают с остатками растений и животных. Жиры рыб, морских животных и растительных масел характеризуются большим количеством ненасыщенных жирных кислот и поэтому подвергаются быстрому окислению. Многие микроорганизмы имеют фермент липазу, катализирующую гидролиз жиров до глицерина и жирных кислот. Большинство микроорганизмов легко осуществляют этот процесс, но с трудом разрушают высшие жирные кислоты. Дальнейшее окисление глицерина и жирных кислот идет различными путями.
Глицерин фосфорилируется до фосфоглицериновой кислоты. Затем фосфоглицериновая кислота окисляется до фосфоглицеринового альдегида. Фосфоглицериновый альдегид гликолитическим путем превращается в пировиноградную кислоту. Пируват в зависимости от видовой специфичности микроорганизмов подвергается дальнейшим превращениям. Жирные кислотыокисляются медленно. Превращение жирных кислот состоит из ряда β-окислений с участием ферментов липооксигеназ. Уксусная кислота (получающаяся при β-окислении жирных кислот) подвергается дальнейшим превращениям через ряд промежуточных реакций цикла Кребса до СО2 и воды. Жирные кислоты окисляются с последовательным отделением двух атомов углерода, дающих уксусную кислоту, и с образованием новой жирной кислоты. Новая жирная кислота подвергается снова β-окислению. Это окисление происходит до образования оксикислот, затем до образования кетокислот. Кетокислоты подвергаются декарбоксилированию с образованием алкилметилкетонов (рисунок Г.5). Кроме того, в окисленных жирах встречаются альдегиды, моно- и диглицериды, вторичные спирты, лактоны.
Липолитические микроорганизмы распространены в почве, воде, воздухе, на продуктах растительного и животного происхождения. Выраженной липолитической активностью обладают бактерии Pseudomonas fluorescens, Achromobacter lipoliticum, представители рода Micrococcus, Streptococcus aureus(особенно активны они в период интенсивного роста), грибы родов Aspergillus, Penicillium, Oidium lactis (молочная плесень), некоторые дрожжи.
Разложение липопротеидов микроорганизмами сопровождается отщеплением липидов с их последующим распадом. Например, холин (составная часть лецитина) в процессе гниения превращается в триметиламин, диметиламин и метиламин. При окислении триметиламина образуется оксид триметиламина, имеющий рыбный запах:
СН2ОН-СН2-ОНN(СН3)3 → N(СН3)3 → О=N(СН3)3
Из холина при гниении может также образовываться ядовитое вещество нейрин. В анаэробных условиях образуется фосфористый водород (фосфин), обладающий неприятным запахом.
§
13 Уксусную кислоту можно получать следующими способами:
1) Из метана, являющегося основным компонентом природного газа, получают ацетилен:

Ацетилен в присутствии сульфата ртути (II) присоединяет воду, образуется уксусный альдегид (реакция Кучерова):
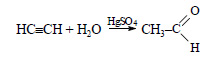
При окислении уксусного альдегида образуется уксусная кислота:

2) При гидратации этилена, являющегося побочным продуктом крекинга нефти, получают этиловый спирт:

При окислении этилового спирта получают уксусную кислоту:

3) Уксусную кислоту можно получить при окислении бутана в присутствии катализатора:

Наиболее перспективным является последний метод, так как бутан является компонентом природного газа и попутных нефтяных газов, а уксусная кислота получается в одну стадию.
Как одна из простейших органических кислот, уксусная кислота является естественным метаболитом (промежуточным продуктом жизнедеятельности) очень многих живых организмов. В естественной среде, например, она является типичным промежуточным звеном концевого окисления этилового спирта CH3-CH2-OH (его формула в ином представлении — C2H5OH), что и определило её первичный способ получения, известный человеку — сбраживание сахаров (прежде всего глюкозы, фруктозы и их производных) напрямую либо через промежуточные стадии спирта этилового и альдегида уксусного (CH3-CHO). В зависимости от происхождения используемого сырья/способа производства, к слову «уксус» нередко добавляется соответствующее прилагательное: «винный уксус», «рисовый уксус», «яблочный уксус» и так далее — и все эти вещества издревле использовались при приготовлении пищи и консервировании/обработке разнообразных пищевых/непищевых продуктов.
Помимо использования в быту и пищевой индустрии (уксусная кислота является официально зарегистрированной пищевой добавкой/консерв
антом с кодом E-260), она также имеет большое промышленное значение — и как компонент моющих/чистящих средств, продуктов гигиены и дезинфекции, и как промежуточное сырьё либо технический продукт (например, специфический растворитель в целом ряде промышленных синтезов). Поскольку массовое производство технической уксусной кислоты «естественным» путём микробиологической переработки пищевых продуктов дорого/неэффективно и имеет смысл лишь в случае наличия некоторых типов бросового либо условно-бесплатного сырья, постоянно изыскивались способы иного её получения.
Образование лимонной кислоты
Лимонная кислота содержится в различных растениях. Она была впервые выделена К. В. Шееле из лимонного сока, где её содержание достигает 10%. В промышленности лимонную кислоту получают путем лимоннокислого брожения отходов сахарного производства с помощью плесневого грибка Aspergillius niger
Применение лимонной кислоты
Лимонная кислота нашла широкое применение в пищевой промышленности (производство плавленых сыров, напитков и т.д.), при производстве косметики, цемента, а также в нефтехиии.
Соли лимонной кислоты называются цитратами. Цитрат натрия применяется для консервации донорской крови. Противосвертывающее действие основано на том, что цитрат натрия связывает участвующие в процессе свертывания ионы кальция в нерастворимый цитрат кальция.
§
Молочную кислоту открыл шведский химик Карл Шееле. В 1780 году он выделил её из прокисшего молока в виде коричневого сиропа. Французский химик Анри Браконно обнаружил, что она образуется при молочнокислом брожении[1].
В 1807 году Йенс Якоб Берцелиус выделил из мышц цинковую соль молочной кислоты.
Молочная кислота – это органическое соединение с формулой CH3CH(OH)CO2H. Визуально это белое, твердое, растворимое в воде вещество или прозрачная жидкость, которая может быть натуральной или вырабатываться синтетическим путем. Молочная кислота (пищевая добавка Е270) состоит из двух оптических изомеров. Один из них известен как L-( )-молочная кислота, а другой, его зеркальное отражение, D-(-)-молочная кислота. Их смесь в равных количествах называется DL-молочная кислота.
В 1856 году Луи Пастер обнаружил лактобактерии и выяснил их роль в создании молочной кислоты. Эти бактерии способны также размножаться во рту. Кислота, которую они производят, приводит к заболеванию зубов, которое известно как кариес. В промышленности молочнокислое брожение осуществляется с помощью молочнокислых бактерий, которые превращают простые углеводы, такие как глюкоза, сахароза, галактоза, в молочную кислоту. Источниками углеводов являются кукуруза, свекла и тростниковый сахар. Молочная кислота также производится путем химического синтеза.
Пищевая добавка Е270 используется в качестве пищевого консерванта, регулятора кислотности и ароматизатора. Этот ингредиент в обработанных пищевых продуктах также используется при обработке мяса в качестве дезинфицирующего средства. Данное вещество также используется для повышения стабильности картофельных продуктов.
Естественная молочная кислота содержится преимущественно в кисломолочных продуктах, таких как кумыс, йогурт, кефир, некоторые сыры и чайный гриб. Казеин в кисломолочных продуктах сгущается с помощью молочной кислоты, которая также отвечает за кислый вкус закваски хлеба. Данное вещество содержится в таких продуктах, как конфеты, заправки для салатов, торты, печенье, готовое к употреблению мясо и птица, пиво, кислое молоко и йогурт, безалкогольные напитки, детские смеси, кондитерские изделия, маринованные продукты, консервированные фрукты и овощи. Молочная кислота также применяется в фармацевтике для производства водорастворимых лактатов из нерастворимых активных ингредиентов. Они применяются в средствах ухода за кожей и косметике для регулирования кислотности и обладают дезинфицирующими и кератолитическими свойствами.
Микроорганизмы, осуществляющие трансформацию органических веществ
Современная методология микробной трансформации позволяет использовать для осуществления того или иного химического превращения в принципе любой микроорганизм, имеющий соответствующие ферменты. Требования, предъявляемые к микробному штамму, пригодному для использования в исследовательской и производственной практике, сводятся к следующему:
1) микроорганизм должен развиваться на сравнительно простых средах;
2) активность фермента и ферментной системы, ответственных за трансформацию, должна быть достаточна высокой;
3) накопление продукта трансформации в среде должно быть достигнуто наиболее простыми методами;
4) перечисленные выше условия должны обеспечивать экономическую рентабельность процесса.
В соответствии с этими требованиями для микробной трансформации органических соединений используются обычно сапрофитные микроорганизмы, способные расти на обычных микробиологических средах и отличающиеся интенсивным обменом веществ. Набор микробов, применяемых в настоящее время в лабораторной и производственной практике для препаративного получения органических веществ методом микробной трансформации, очень широк. Он включает представителей грибов (аскомицеты, фикомицеты, базидиомицеты, несовершенные грибы), актиномицетов и родственных им организмов, многих других бактерий и даже микроформы водорослей.
Попытки многих исследователей установить видовую и родовую специфичность микроорганизмов, осуществляющих различные трансформации, не всегда приводили к успеху. Можно сказать с определенностью, что, например, для гидроксилирования алкильных заместителей ароматических соединений следует искать активные штаммы в группе rhodochrousрода Nocardia,для окисления оксигрупп полиолов — среди уксуснокислых бактерий, для изомеризации альдоз — среди стрептомицетов бурой группы, артробактеров, лактобацилл, бацилл и др. Однако в большинстве случаев приходится ориентироваться на более крупные таксоны и более широкий поиск, что усложняет задачу.
Таким образом, потенциальная способность осуществлять различные трансформационные процессы распространена весьма широко среди микроорганизмов, и далеко не всегда можно заранее указать узкую таксономическую группу, в которой следует искать штаммы, осуществляющие определенное превращение.
Существуют, однако, некоторые таксоны — роды и даже виды, способные проводить разнообразные превращения органических веществ с накоплением продуктов трансформации. Среди них заслуживают особого внимания G. oxydans, осуществляющий десятки различных окислительных превращений углеводов и родственных соединений, Aspergillus niger, который наряду с окислением углеводов используется также для их восстановления, гидролиза гликозидных связей и трансгликозидирования, Brevibacterium ammoniagenes, который синтезирует рибозиды и нуклеотиды из предшественников и аминирует органические кислоты. Очень часто упоминаются в литературе проводящие различные трансформации представители родов Arthrobacter, Corynebacterium, Mycobacterrium, Pseudomonas.
§
Процесс хемосинтеза в биологии представляет собой в некотором смысле уникальное явление, ведь это необычный тип питания бактерий, основанный на усвоении углекислого газа СО2благодаря окислению неорганических соединений. Причем что интересно, по мнению ученых, хемосинтез это древнейший тип автотрофного питания (такого питания, когда организм сам синтезирует органические вещества из неорганических), который мог появиться даже раньше нежели фотосинтез.
История открытия хемосинтеза
Как биологическое явление хемосинтез бактерий был открыт русским биологом С. Н. Виноградским в 1888 году. Ученый доказал способность некоторых бактерий выделять углеводы используя химическую энергию. Им же был выделен ряд особых хемосинтизирующих бактерий, среди которых наиболее заметными являются серобактерии, железобактерии и нитрифицирующие бактерии.
Хемосинтез и фотосинтез: сходства и различия
Давайте теперь разберем в чем сходство хемосинтеза и фотосинтеза, а в чем различия между ними.
Сходство:
§ Как хемосинтез, так и фотосинтез являются типами автотрофного питания, когда организм выделяет органические вещества из неорганических.
§ Энергия такой реакции запасается в аденозинтрифосфорной кислоте (сокращено АТФ) и впоследствии используется для синтеза органических веществ.
Отличие фотосинтеза от хемосинтеза:
§ У них разный источник энергии, и как следствие разные окислительно-восстановительных реакции. При хемосинтезе первичным источником энергии является не солнечный свет, а химические реакции по окислению определенных веществ.
§ Хемосинтез характерен исключительно для бактерий и арей.
§ При хемосинтезе клетки бактерий не содержат хлорофилла, при фотосинтезе наоборот – содержат.
§ Источником углерода для синтеза органики при хемосинтезе может быть не только лишь углекислый газ, но и окись углерода (СО), муравьиная кислота, уксусная кислота, метанол и карбонаты.
Энергия хемосинтеза
Свою энергию бактерии хемосинтетики получают благодаря окислению водорода, марганца, железа, серы, аммиака и т. д. В зависимости от окисляемого субстрата упомянутые нами выше бактерии и получили свои названия: железобактерии, серобактерии, метанобразующие археи, нитрифицирующие бактерии, ну и так далее.
Значение хемосинтеза в природе
Хемотрофы – организмы, получающие жизненную энергию благодаря хемосинтезу, играют важную роль в круговороте веществ, особенно азота, в частности они поддерживают плодородность почв. Также благодаря деятельности бактерий-хемосинтетиков в природных условиях накапливаются большие запасы руды и селитры.
Реакции хемосинтеза
Теперь давайте более детально разберем существующие реакции хемосинтеза, все они отличаются в зависимости от бактерий-хемосинтетиков.
Железобактерии
К ним относятся нитчатые и железоокисляющие лептотриксы, сферотиллюсы, галлионеллы, металлогениумы. Обитают они в пресных и морских водоемах. Благодаря реакции хемосинтеза образуют отложения железных руд путем окисления двухвалентного железа в трехвалентное.
4FeCO3 O2 6H2O → Fe(OH)3 4CO2 E (энергия)
Помимо энергии в этой реакции образуется углекислый газ. Также помимо бактерий окисляющих железо, есть бактерии окисляющие марганец.
Серобактерии
Иное их название – тиобактерии, представляют собой весьма большую группу микроорганизмов. Как это следует из их названия, эти бактерии получают энергию путем окисления соединений с восстановленной серой.
2S 3O2 2H2O → 2H2SO4 E
Полученная в результате реакции сера может, как накапливаться в самих бактериях, так и выделятся в окружающую среду в виде хлопьев.
Нитрифицирующие бактерии
Эти бактерии, обитающие в земле и воде, свою энергию получают за счет аммиака и азотистой кислоты, именно они играют очень важную роль в кругообороте азота.
2NH3 3O2 → HNO2 2H2O E
Азотистая кислота, полученная при такой реакции, образует в земле соли и нитраты, способствующие ее плодородию.
Нитрификация и микроорганизмы ее осуществляющие.
Нитрификация — микробиологический процесс окисления аммиака до азотистой кислоты или её самой далее до азотной кислоты, что связано либо с получением энергии (хемосинтез, автотрофная нитрификация), либо с защитой от активных форм кислорода, образующихся при разложении пероксидаводорода (гетеротрофная нитрификация).
Протекает в аэробных условиях в почве, а также природных водах. Часто может вызывать появление в них нитратов в токсичном количестве, а поскольку нитраты — наиболее активно мигрирующее в растворе соединение азота — происходит их вынос из почвы в расположенные ниже по склону водоемы, что влечет за собой уменьшение коэффициента использования азотных удобрений и эвтрофикацию данных водоемов.
Нитрификация проходит в две стадии, которые осуществляются разными микроорганизмами.
§
Первая стадия — окисление аммиака до нитрит-аниона, которое осуществляют нитрозные бактерии родов Nitrosomonas, Nitrosococcus и Nitrosospira (ранее выделялись также рода Nitrosolobus, Nitrosovibrio, но сейчас их представители включены в Nitrosospira) по следующему механизму:
1. NH3 O2 НАДН2 → NH2OH H2O НАД
2. NH2OH H2O → HNO2 4H 4e−
3. 1/2O2 2H 2e− → H2O
Предполагается, что на первом этапе субстратом является именно аммиак, а не аммоний, поэтому процесс не идёт в кислой среде. Ферментом для первой реакции служит аммиакмонооксигеназа, фермент с очень низкой субстратной специфичностью, окисляющая также метан, оксид углерода, циклогексан, фенол, бензиловый спирт, однако со скоростью на порядки ниже. Гидроксиламин ингибирует работу фермента. В бесклеточных экстрактах восстановителем может служить НАД(Ф)·H, однако в клетке его роль, скорее всего, выполняет один из компонентов дыхательной цепи.
Следующую реакцию осуществляет гидроксиламиноксидоредуктаза, расположенная в периплазме. Окислителем в них служит цитохром c, с него электрон передаётся на убихинон и далее в дыхательную цепь, на цитохромоксидоредуктазу и, в конечном итоге, на кислород. При этом запасается энергия в виде трансмембранного протонного потенциала.
Образование НАД(Ф)·H для фиксации углекислого газа в цикле Калвина происходит путём обратного переноса части электронов.
Окисление нитрит-аниона до нитрат-аниона[править | править код]
Вторая стадия — окисление аниона азотистой кислоты до аниона азотной, производимое нитратными бактериями (почвенный род Nitrobacter и водные Nitrospira, Nitrococcus, Nitrospina). Процесс протекает в одну реакцию:
NO2− H2O → NO3− 2H 2e−
катализируемую нитрит: нитрат-оксидоредуктазой, локализованной в ЦПМ. Далее электроны передаются на цитохромы дыхательной цепи, в которой единственным пунктом транслокации протонов является цитохромоксидаза.
Образование НАД(Ф)·H для фиксации углекислого газа также происходит путём обратного переноса электронов.
Организмы[править | править код]
Нитрозных и нитритных бактерий ранее выделяли в семейство Nitrobacteraceae, сейчас с развитием геносистематики их рода разнесены по разным подклассам протеобактерий. Оптимальная для их развития температура 25—30 °C и pH 7,5—8,0. В кислой среде процесс не идет. Все эти бактерии — грамотрицательные хемолитоавтотрофы, использующие энергию окисления соединений азота для синтеза органических веществ из углекислого газа. Некоторые из них способные переключаться на хемоорганогетеротрофный метаболизм, однако ни одного хемолитогетеротрофа среди данных организмов найдено не было. Морфологически эти группы разнообразны, в большинстве своем мелкие, подвижные, с полярным или перитрихиальным жгутикованием. Окисление они проводят на цитоплазматической мембране.
Гетеротрофная нитрификация[править | править код]
Особо выделяют гетеротрофную нитрификацию, происходящую у многих бактерий и грибов и связанную с окислением аммиака и азота органических соединений без использования полученной энергии, попутно с окислением органического вещества и, предположительно, посредством кислорода, образуемого при разложении пероксида водорода. Удельная активность этого процесса на 2-4 порядка ниже, чем в случае автотрофной нитрификации, однако именно с ним связано происхождение чилийской селитры а также вся нитрификация в почвах и водоемах с низкими значениями pH.
История изучения[править | править код]
Процесс был открыт в середине XIX века, первое предположение об участии микроорганизмов в нитрификации было сделано Луи Пастером, однако выделить их не удавалось, пока в 1891 С. Н. Виноградский не применил для этих целей селективную среду, содержащую только минеральные компоненты (чтобы избежать роста на ней банальных гетеротрофов) и в том числе сернокислый аммоний.
Аммиак, образующийся при аэробном и анаэробном разложении азотсодержащих органических веществ, окисляется двумя высокоспециализированными группами микроорганизмов в азотистую и затем азотную кислоты.
Этот процесс называется нитрификацией. Первыми исследователями, доказавшими его биологическую природу, были Т. Шлезинг и А. Мюнц (1879). На специфичность организмов, вызывающих нитрификацию» указывал Луи Пастер (1880). Однако длительное время их не удавалось выделить в чистую культуру. Лишь в 1890—1892 гг. С. Н. Виноградскому, применившему элективные среды, не содержащие органические вещества, удалось изолировать чистые культуры нитрификаторов. Им было установлено и существование двух групп нитрификатов, одна из которых окисляет аммиак до азотистой кислоты, а другая — азотистую кислоту в азотную. Последовательность в работе — двух или нескольких организмов называется метабиозом.
Нитрифицирующие бактерии входят в семейство Nitrobacteriaceae, имеют ряд общих свойств: все — облигатные аэробы, грамотрицательны, не образуют спор, развиваются при pH 7,5—8,0, оптимальная температура для их роста 25—30° С. Однако различные их роды существенно морфологически отличаются.
Первый этап нитрификации осуществляют бактерии пяти родов: Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosovibrio, Nitrosolobus; наиболее изучен род Nitrosomonas. В целях получения энергии для ассимиляции 1,5 г СO2 и построения 1 г микробной плазмы нитрозные бактерии окисляют около 20 г аммиака и образуют 56 г азотистой кислоты; реакция: 2NH3 3O2 = 2HNO2 2H2O 158 кал. Окисление аммиака совершается в несколько стадий: на поверхности клетки — до гидроксиламина, далее уже внутри клетки — до гипонитрита и окиси азота.
Микроорганизмы, способные окислять нитриты в нитраты (осуществляющие вторую фазу нитрификации), относятся к родам Nitrobacter, Nitrococcus, Nitrospina. Детально исследован род Nitrobacter. Все его представители имеют клетки грушевидной формы, размножаются почкованием, у дочерней клетки есть жгутик, оптимум развития при pH 7,7. Окисление азотистой кислоты в азотную идет также с выделением энергии: 2HNO2 O2 -> 2HNO3 48 кал. При построении 1 г вновь синтезированного органического вещества микробных тел окисляется 72 г HNO2 и образуется 96 г HNO3. Роль нитрификаторов в образовании нитратов грандиозна.
Воздействие органического вещества на нитрификаторы двояко: в лабораторных условиях даже ничтожные дозы органического вещества полностью подавляют рост и развитие организма, а в естественных условиях — нет. Причины этого различия не вполне ясны.
Наряду с нитрифицирующими автотрофными бактериями Д. У. Катлер и Б. К. Мукерджи (1931) выделили ряд гетеротрофных бактерий, способных окислять NH4 до NO2, но значительно медленнее и только на первом этапе. В настоящее время установлены микроорганизмы, доводящие нитрификацию до конца, в частности гриб Aspergillus flavus и некоторые виды Atrobacter, нитрифицирующие аммиак.
К гетеротрофным нитрификаторам относят представителей родов: Alcaligenes, Corynebacterium, Achromobacter, Pseudomonas, Nocardia, Bacillus. Многие гетеротрофы могут осуществлять нитрификацию органических азотсодержащих соединений: амидов, аминов, оксидов, гидроксаматов и других, а также образовывать продукты, обладающие канцерогенными, токсичными, мутагенными свойствами. В условиях, неблагоприятных для развития автотрофных нитрифицирующих бактерий нитрификаторы-гетеротрофы, по-видимому, главенствуют в нитрификационном процессе.
§
Каждый микроорганизм может развиваться лишь в определенных пределах температуры. При изучении влияния температуры на рост прокариот выделяют температурный диапазон, ограниченный минимальной и максимальной температурами, при которых рост прекращается, а также область оптимальных температур с максимальной скоростью роста.
Положение на температурной шкале основных точек (минимальная, максимальная, оптимальная температуры), а также величина температурного диапазона роста прокариот сильно различаются.Поэтому по отношению к температуре микроорг-змы подразд-ют на 3 группы: мезофилы, психрофилы и термофилы.
(У мезофилов оптимальные температуры роста лежат между 30 и 40 °С, а температурный диапазон, в котором возможен рост, находится между 10 и 45-50 °С (например Escherichia coli: нижняя граница роста — 100С, верхняя — 49 °С, оптимальная температура — 37 °С при росте на богатой среде.)
У психрофилов (холодолюбивые микроорганизмы) область температур роста лежит в пределах от —10 до 20 °С и выше.Они делятся на облигатных (обязательных) и факультативных (необязательных), (основное различие между ними в том, что облигатные психрофилы не способны к росту при температуре выше 20 °С, а верхняя температурная граница роста факультативных форм намного выше. Таким образом, факультативные психрофилы характеризуются более широким температурным диапазоном, при котором возможен их рост.)
Термофилов (теплолюбивые микроорганизмы) делят на 4 подгруппы.
1. Термотолерантные виды растут в пределах от 10 до 55—60 °С, оптимальная область лежит в пределах 35—40 °С. (Они способны расти при повышенных температурах.)
2. Факультативные термофилы имеют максимальную температуру роста между 50 и 165 0С, но способны к размножению при температуре 20 °С; оптимум приходится на область температур, близких к верхней границе роста. (Особенность этой группы прокариот — способность к росту в области от 20 до 40 °С.)
3. К облигатным термофилам относят виды, обнаруживающие способность расти при температурах около 70 °С и не растущие ниже 40 0С. (Оптимальная температурная область облигатных термофилов примыкает к их верхней температурной границе роста. Представители: эубактерии Bacillusacidocaldarius, Synechococcus lividus, архебактерии Methanobacterium thermoautotrophicum, Thermo- plasma acidophilum и др.)
4. Экстремальные термофилы для них характерны: оптимальная граница роста — в области 80— 1050С, минимальная — при 60 °С и выше, максимальная — до 1100С. (Thermoproteus, Pyrococcus, Pyrodictium и др.))
Наиболее термоустойчивыми являются бактериальные споры. У многих бактерий они способны выдерживать температуру кипения воды в течение нескольких часов. Во влажной среде споры бактерий гибнут при 120—130 °С через 20—30 мин, в сухом состоянии — при 160—170 °С через 1—2 ч. Термоустойчивость спор различных бактерий неодинакова; особенно устойчивы споры термофильных бактерий.
Механизм действия температуры на микроорганизмы. Отрицательное влияние на жизнедеятельность микроорганизмов оказывает и низкая и высокая температура. Из-за низкой температуры замедляются биохимические процессы и в случае образования кристаллов льда внутри клетки происходит повреждение и разрыв клеточных структур. Низкая температура вызывает бактериостатинеский эффект, который проявляется в прекращении роста и размножения бактерий.
Один из лучших методов длительного хранения скоропортящихся продуктов с минимальными изменениями их химического состава – консервирование низкими температурами. Низкие температуры замедляют химические и биохимические процессы обмена веществ в тканях, снижают ферментативную активность, приостанавливают развитие микроорганизмов. Консервирование низкими температурами проводят путем охлаждения или замораживания.
Охлаждением называется обработка и хранение пищевых продуктов при температуре, близкой к криоскопической, т. е. к температуре замерзания клеточного сока, которая зависит от состава и концентрации сухих веществ. (Для яблок она колеблется от —1,4 до —2,8 °С, для винограда равна —3,8 °С, для лука —1,6, для рыбы —2, для мяса — 1,2 °С и ниже.) (При хранении охлажденных продуктов лучше, чем при замораживании, сохраняются их натуральные свойства, но рост на них многих микроорганизмов не исключается, а лишь замедляется, поэтому сроки хранения охлажденных продуктов непродолжительны и зависят от температуры хранения и исходной степени обсеменения продукта психрофильными микроорганизмами.)
При замораживании происходит полная кристаллизация жидкой фазы продукта. Этот способ применяется для более длительного сохранения мясных и рыбных продуктов, овощей, фруктов и др. (При длительном хранении замороженных продуктов изменяется их химический состав, гидролизуются и окисляются жиры, изменяется цвет, частично разрушаются витамины в результате их окисления кислородом воздуха, ухудшается вкус и запах. При замораживании микрофлора полностью не уничтожается; особенно холодоустойчивы споровые формы микроорганизмов. Во время размораживания продуктов, особенно при вытекании из них сока, микроорганизмы вновь размножаются и вызывают порчу. С учетом этого оттаивать замороженные пищевые продукты следует непосредственно перед употреблением. Кроме того, после размораживания продукта микроорганизмы возобновляют свою жизнедеятельность и могут привести к его быстрой порче, поэтому размороженные продукты необходимо сразу же перерабатывать. Качество продуктов в значительной степени зависит от способа его размораживания. При быстром размораживании при повышенных температурах происходят большие потери питательных веществ и более интенсивно развиваются микроорганизмы. Во время медленного размораживания при температурах от 0 до 4 °С кристаллы льда оттаивают постепенно, а коллоиды клеток более полно связывают образующуюся влагу. )
Наиболее губительна для микроорганизмов высокая температура, при которой происходит свертывание белка, вследствии чего происх-т нарушение активности ферментов, проницаемости клеточной стенки и в рез-те чего нарушается равновесие всех биохимических процессов. Сущ-ют след. приемы уничтожения микробов в пищевых продуктах при высоких температурах: варка, кипячение, обжарка, бланширование продуктов питания, пропаривание производственного оборудования. В пищевой промышл-сти применяют 2 способа воздействия высоких температур на микроорганизмы: пастеризацию и стерилизацию.
Пастеризация — это нагревание продукта чаще при температуре 63—80 °С в течение 20-40 мин. Иногда пастеризацию производят кратковременным (в течение нескольких секунд) нагреванием до 90—1000С. При пастеризации погибают не все микроорганизмы. Поэтому пастеризованные продукты необходимо немедленно охлаждать до температуры не выше 10 “С и хранить на холоде, чтобы задержать прорастание спор и развитие сохранившихся клеток. Пастеризуют молоко, вино, соки, икру, , варенье, плодово-ягодные компоты и другие продукты. Иногда для удлинения сроков хранения продуктов проводят многократную пастеризацию — тиндализацию.
Стерилизация — это нагревание при температурах, которые вызывают гибель всех живых микроорганизмов. В микробиологической практике стерилизуют среды, посуду, инструмент и другие необходимые предметы. Существуют различные способы стерилизации при помощи высокой температуры.
Асептическое консервирование состоит в том, что жидкие и пюреобразные пищевые продукты подвергают стерилизации путем кратковременного высокотемпературного нагрева, охлаждают, а затем расфасовывают в стерильную тару и укупоривают в асептических условиях ( применяют для консервирования томата-пасты, плодово-ягодных соков, молока и других продуктов. Преимущество такого способа состоит в том, что сокращается время тепловой обработки продукта, в результате чего повышается пищевая ценность консервов, для упаковки могут быть использованы полимерные материалы.)
§
Развитие всех микроорганизмов возможно при определенной температуре. Известны микроорганизмы, способные существовать при низких (-8°С и ниже) и при повышенных температурных условиях, например, обитатели горячих источников поддерживают жизнедеятельность при температуре 80-95°С. Большинство микробов предпочитает температурные пределы 15-35°С. Различают:
· оптимальную, наиболее благоприятную для развития температуру;
· максимальную, при которой прекращается развитие микробов данного вида;
· минимальную, ниже которой микробы прекращают развитие.
По отношению к уровню температуры микроорганизмы разделяют на три группы:
· психрофиты – хорошо растут при пониженных температурах,
· мезофиллы – нормально существуют при средних температурах,
· термофилы – существуют при постоянно высоких температурах.
Группа микроорганизмов | Температура развития микроорганизмов, ° С | ||
| Минимальная | Оптимальная | Максимальная | |
| Психрофилы | 0-2 | 15-25 | 25-35 |
| Мезофилы | 25-37 | 40-45 | |
| Термофилы | 45-60 | ||
Микробы сравнительно быстро приспосабливаются к значительным изменениям температуры. Поэтому незначительное снижение или повышение уровня температуры не гарантирует прекращения развития микроорганизмов.
Влияние высоких температур.
Температуры, значительно превышающие максимальные, вызывают гибель микроорганизмов. В воде большинство вегетативных форм бактерий при нагревании до 60°С погибают за час; до 70°С — за 10-15 минут, до 100°С — за несколько секунд. В воздухе гибель микроорганизмов наступает при значительно более высокой температуре — до 170°С и выше в течение 1-2 часов. Споровые формы бактерий значительно устойчивее к нагреванию, они могут выдерживать кипячение в течение 4-5 часов.
Методы пастеризации и стерилизации основаны на свойстве микробов погибать под действием высоких температур. Пастеризация — осуществляется при температуре 60-90°С, при этом погибают вегетативные формы клеток, а споровые остаются жизнеспособными. Поэтому пастеризованные продукты следует быстро охлаждать и хранить в условиях охлаждения. Стерилизация — это полное уничтожение всех форм микроорганизмов, включая споровые. Стерилизацию осуществляют при температуре 110-120°С и повышенном давлении.
Однако споры не погибают мгновенно. Даже при 120°С гибель их наступает через 20-30 минут. Стерилизуют пищевые консервы, некоторые медицинские материалы, субстраты, на которых выращивают микроорганизмы в лабораториях. Эффект стерилизации зависит от количественного и качественного состава микрофлоры объекта стерилизации, его химического состава, консистенции, объема, массы и др.
Влияние низких температур.
Чаще всего действие низких температур связано не с гибелью микроорганизмов, а с торможением и прекращением их развития. Низкую температуру микроорганизмы переносят значительно лучше. Многие болезнетворные микробы, попадающие в окружающую среду, способны переносить суровые зимы, не теряя болезнетворности. Наиболее негативно на развитие микроорганизмов влияет температура, при которой замерзает содержимое клетки.
Тормозящее действие низких температур на микробы используют для хранения различных продуктов в охлажденном виде при температуре 0-4°С, и замороженном – при температуре — 6-20°С и ниже. Действие низких температур в замороженных продуктах усиливает влияние повышенного осмотического давления. Поскольку большая часть воды перешла в лед, в оставшейся жидкой части воды оказались все растворенные вещества, содержавшиеся в массе продукта. Это вызывает повышенное осмотическое давление, которое, в свою очередь, тормозит развитие микробов.
Замораживание используют для хранения мяса, рыбы, плодов, овощей полуфабрикатов, кулинарных изделий, готовых блюд и др. Прекращение развития микробов действует только до тех пор, пока продолжается действие низкой температуры. При повышении температуры начинается бурное развитие и размножение микробов, что вызывает порчу пищевых продуктов.
Следовательно, низкая температура только замедляет биохимические процессы, не имея стерилизующего эффекта. Многократное замораживание одних и тех же продуктов способствует быстрому приспособлению микробов к низким температурам и усиливает их жизнеспособность. Поэтому надо предотвращать колебания температуры во время хранения продуктов.
§
Бактериальные организмы могут существовать и развиваться в весьма широких температурных границах.
Различают три экологические группы бактерий по их отношению к температуре окружающей среды: психрофилы, мезофилы, термофилы.
К психрофилам относятся формы, развивающиеся при сравнительно низкой температуре. Оптимум развития у этих бактерий лежит при 20—25 °C, а минимум может лежать даже ниже нуля, если эти бактерии развиваются в растворе, замерзающем при температуре ниже нуля.
К мезофильным бактериям относится огромное большинство бактерий. Оптимум развития мезофилов лежит при 25—35 °C, максимум — при 45—50 °C и минимум — около 10 °C. Из приведенных для мезофилов температурных границ видно, что эта группа микроорганизмов наиболее соответствует температурам, наблюдающимся в природе в летнее время, и поэтому не удивительно весьма широкое распространение мезофилов в почве, воде и других субстратах.
К термофильным микроорганизмам относятся бактерии и близкие к ним актиномицеты, а также некоторые водоросли, по преимуществу сине-зеленые. Все они характеризуются тем, что развиваются при высоких температурах (40—75 °C). Водоросли и ряд представителей животного мира (моллюски и др.), живущие в горячих источниках и гейзерах, развиваются при несколько более низких температурах, чем бактерии и актиномицеты. Из микроорганизмов к термофилам обычно относятся бациллярные формы, образующие споры, а также группа актиномицетов.
В состоянии спор термофилы хорошо переносят температуры ниже 30 °C, при которых они в вегетирующем состоянии быстро гибнут. Особенностью термофилов является их очень интенсивный обмен, благодаря чему они гораздо энергичнее размножаются и значительно быстрее заканчивают осуществляемые ими микробиологические процессы. Клетки термофилов делятся быстрее, чем у мезофилов, и быстрее стареют и отмирают.
Влияние температуры на развитие микроорганизмов
Развитие всех микроорганизмов возможно при определенной температуре. Известны микроорганизмы, способные существовать при низких (-8°С и ниже) и при повышенных температурных условиях, например, обитатели горячих источников поддерживают жизнедеятельность при температуре 80-95°С. Большинство микробов предпочитает температурные пределы 15-35°С. Различают:
· оптимальную, наиболее благоприятную для развития температуру;
· максимальную, при которой прекращается развитие микробов данного вида;
· минимальную, ниже которой микробы прекращают развитие.
По отношению к уровню температуры микроорганизмы разделяют на три группы:
· психрофиты – хорошо растут при пониженных температурах,
· мезофиллы – нормально существуют при средних температурах,
· термофилы – существуют при постоянно высоких температурах.
Группа микроорганизмов | Температура развития микроорганизмов, ° С | ||
| Минимальная | Оптимальная | Максимальная | |
| Психрофилы | 0-2 | 15-25 | 25-35 |
| Мезофилы | 25-37 | 40-45 | |
| Термофилы | 45-60 | ||
Микробы сравнительно быстро приспосабливаются к значительным изменениям температуры. Поэтому незначительное снижение или повышение уровня температуры не гарантирует прекращения развития микроорганизмов.
Влияние высоких температур.
Температуры, значительно превышающие максимальные, вызывают гибель микроорганизмов. В воде большинство вегетативных форм бактерий при нагревании до 60°С погибают за час; до 70°С — за 10-15 минут, до 100°С — за несколько секунд. В воздухе гибель микроорганизмов наступает при значительно более высокой температуре — до 170°С и выше в течение 1-2 часов. Споровые формы бактерий значительно устойчивее к нагреванию, они могут выдерживать кипячение в течение 4-5 часов.
Методы пастеризации и стерилизации основаны на свойстве микробов погибать под действием высоких температур.
Пастеризация — осуществляется при температуре 60-90°С, при этом погибают вегетативные формы клеток, а споровые остаются жизнеспособными. Поэтому пастеризованные продукты следует быстро охлаждать и хранить в условиях охлаждения.
Стерилизация — это полное уничтожение всех форм микроорганизмов, включая споровые. Стерилизацию осуществляют при температуре 110-120°С и повышенном давлении.
Однако споры не погибают мгновенно. Даже при 120°С гибель их наступает через 20-30 минут. Стерилизуют пищевые консервы, некоторые медицинские материалы, субстраты, на которых выращивают микроорганизмы в лабораториях. Эффект стерилизации зависит от количественного и качественного состава микрофлоры объекта стерилизации, его химического состава, консистенции, объема, массы и др.
Влияние низких температур.
Чаще всего действие низких температур связано не с гибелью микроорганизмов, а с торможением и прекращением их развития. Низкую температуру микроорганизмы переносят значительно лучше. Многие болезнетворные микробы, попадающие в окружающую среду, способны переносить суровые зимы, не теряя болезнетворности. Наиболее негативно на развитие микроорганизмов влияет температура, при которой замерзает содержимое клетки.
Тормозящее действие низких температур на микробы используют для хранения различных продуктов в охлажденном виде при температуре 0-4°С, и замороженном – при температуре — 6-20°С и ниже. Действие низких температур в замороженных продуктах усиливает влияние повышенного осмотического давления. Поскольку большая часть воды перешла в лед, в оставшейся жидкой части воды оказались все растворенные вещества, содержавшиеся в массе продукта. Это вызывает повышенное осмотическое давление, которое, в свою очередь, тормозит развитие микробов.
Замораживание используют для хранения мяса, рыбы, плодов, овощей полуфабрикатов, кулинарных изделий, готовых блюд и др. Прекращение развития микробов действует только до тех пор, пока продолжается действие низкой температуры. При повышении температуры начинается бурное развитие и размножение микробов, что вызывает порчу пищевых продуктов.
Следовательно, низкая температура только замедляет биохимические процессы, не имея стерилизующего эффекта. Многократное замораживание одних и тех же продуктов способствует быстрому приспособлению микробов к низким температурам и усиливает их жизнеспособность. Поэтому надо предотвращать колебания температуры во время хранения продуктов
§
Пастеризация — однократное
нагревание жидкостей или пищевых продуктов обычно до 60-70°С в течение 15-30 минут. Была открыта в середине XIX века французским микробиологом Луи Пастером. Цель пастеризации — уничтожение болезнетворных микроорганизмов и подавление жизнедеятельности микроорганизмов, вызывающих порчу продуктов. Эффективность пастеризации определяется степенью уничтожения болезнетворной (патогенной) микрофлоры. При пастеризации продукт нагревают до температуры 63-98 °С и выдерживают при этой температуре некоторое время. При такой обработке инактивируются ферменты, погибают вегетативные формы микроорганизмов, но споры остаются в жизнеспособном состоянии и при возникновении благоприятных условий начинают развиваться. Поэтому пастеризованные продукты (молоко, пиво и др.) хранят при пониженных температурах в течение ограниченного периода времени. Пищевая ценность продуктов при пастеризации практически не изменяется. В зависимости от вида и свойств пищевого сырья используют разные режимы пастеризации. Различают длительную (при температуре 63—65°С в течение 30-40 мин), короткую (при температуре 85-90°С в течение 0,5-1 мин) и мгновенную пастеризацию (при температуре 98°С в течение нескольких секунд). Стерилизация — уничтожение всех видов микроорганизмов, включая бактерии и их споры, грибы, вирусы и прионы, находящихся на поверхностях, оборудовании, в пищевых продуктах и лекарствах. Осуществляется термическим, химическим, радиационным, фильтрационным методами. Термическая стерилизация . Как и при пастеризации, в стерилизации существует зависимость между температурой нагрева продукта и продолжительностью выдержки. Иногда применяют дробную стерилизацию, которая заключается в многократном нагреве и охлаждении продукта. Сначала продукт нагревают до 100—150 °C, затем его охлаждают до 35-40 °C и выдерживают при этой температуре 1-2 часа. Это делается для того, чтобы проросли споры. Потом продукт вновь нагревают до 100—110°C, затем опять охлаждают до 35-40 °C и выдерживают 1-2 часа. Так повторяют несколько раз. Эффективность стерилизации характеризуют коэффициентом стерилизующего действия, который представляет собой логарифм отношения начального и конечного количества бактерий в единице объема продукта С = lg(N0/Nк) Биотехнология предст-т собой сов-ть пром-х методов, в кот-х исп-ся живые организмы и биолог-ие процессы для произв-ва разл-х продуктов ( хлебопечение, приготовление вина, пива, сыра, способы обработки кожи и т.д.) Биопромышл-ть,в онове которой лежит биотехнология роизводит кормовые и пищевые белки, аминокислоты, ферменты. К важнейшим процессам биотехнологии отн-ся брожение, микробиологический синтез, термическая обработка. Новые направления физико-химической биологии значительно разширили возможности процессов биотехнологии, особенно в генной и клеточной инженерии. Сегодня биотехнология рассм-ся как наука,возникшая на стыке нескольких биологич-х дисциплин: генетики, вирусологии, микробиологии и растениеводства. Она описывает уникальные возм-ти практического использ-ия рез-в в этой области, выдвигает на передний план НТП.
О тношение микроорганизмов к кислороду: аэробы и анаэробы (облигатные, факультативные).
По своему отношению к кислороду микроорганизмы условно делятся на несколько групп. Облигатные аэробы нуждаются в молекулярном кислороде для окисления питательных веществ и осуществления аэробного дыхания. Это значительная часть микроорганизмов, живущих на различных поверхностях и в верхних слоях почвы и водоемов (например, Micrococcus luteus). При культивировании аэробных микроорганизмов необходимо создать условия доступности молекулярного кислорода, так как его растворимость в воде невелика (при 20оС – 6,2 мл/л). Для этого применяют выращивание в тонком слое среды или обогащение ее кислородом различными способами (перемешивание, барботирование и т.д.).
Среди аэробов выделяют группу микроорганизмов-микроаэрофилов, которым кислород необходим, но в концентрации ниже атмосферной (менее 5%). Такие организмы встречаются на границе анаэробной и аэробной зон в естественных местообитаниях. К ним относятся многие тионовые и железоокисляющие бактерии (в том числе, представители родов Beggiatoa, Galionella), а также пресноводные спириллы (Spirillum volutans).
Факультативные анаэробы способны переключать свой обмен веществ в зависимости от наличия или отсутствия кислорода с аэробного дыхания на анаэробные процессы. Как правило, в присутствии кислорода такие микроорганизмы растут быстрее и накапливают больше биомассы. К факультативным анаэробам относятся Saccharomyces cerevisiae, многие энтеробактерии (E. coli) и бациллы.
Облигатные анаэробы не нуждаются в кислороде для своей жизнедеятельности, более того, в его присутствии происходит угнетение или гибель клеток. В эту группу входят метанобразующие археи, гомоацетогенные бактерии, большинство сульфатредукторов, некоторые грибы и простейшие. При культивировании облигатных анаэробов используют специальные приемы. Кислород удаляют из среды кипячением, сосуды тщательно укупоривают резиновыми пробками с металлическими колпачками, газовую фазу культивационного сосуда заменяют на азот или аргон. Для удаления следовых количеств кислорода в среду вносят восстановители (сульфид натрия) и в дальнейшем поддерживают анаэробные условия, исключая попадание кислорода при пересевах.
Группа аэротолерантных анаэробов также не использует кислород в метаболизме, однако некоторое количество кислорода в среде не влияет на их жизнедеятельность. В эту группу входят молочнокислые бактерии и некоторые патогенные стрептококки (Streptococcus pyogenes).
§
Возможность жизни на нашей планете определяется непрерывно протекающим круговоротом основных биогенных элементов (углерода, кислорода, водорода, азота, фосфора, серы и др.). Ведущая роль в процессах трансформации этих элементов принадлежит прокариотам. Приведем характерный пример. Содержание углекислого газа в атмосфере минимально (составляет всего 0,03%), и если бы не происходил постоянный возврат СО2 в атмосферу, этот газ был бы израсходован в процессе фотосинтеза за какие-нибудь 7–40 лет. Дальнейшая жизнь оказалась бы невозможна. Однако этого не происходит. В результате разложения органических соединений различными группами микроорганизмов в атмосферу возвращается 90% углекислого газа, остальные 10% СО2 пополняются в атмосфере за счёт дыхания эукариот, а также за счет хозяйственной деятельности человека.
Помимо углекислого газа, при разложении органических соединений микроорганизмы возвращают в атмосферу и другие газообразные продукты, такие, как Н2, Н2S, N2, СН4. Таким образом, они осуществляют не только деструкцию растительного и животного опада, выполняя роль санитаров планеты, но одновременно регулируют газовый состав атмосферы.
Ведущая роль прокариот в процессах трансформации элементов в биосфере определяется прежде всего огромной численностью микроорганизмов, повсеместным распространением их, а также универсальностью ферментативного аппарата микробной клетки, способной перерабатывать любые вещества субстрата.
Запасы азота в природе очень велики. Он входит в состав всех организмов на Земле. Общее содержание его в организмах составляет более 25 млрд. тонн, большое количество азота находится также в почве. Но еще более грандиозен запас азота в атмосфере: над каждым гектаром почвы поднимается столб воздуха, содержащий около 80000 тонн молекулярного азота. Ежегодно на образование вновь вырастающих растений требуется около 1,5 млрд. тонн азота в форме, доступной для усвоения растениями. Имеющегося в воздухе и почве азота хватило бы для обеспечения урожая, даже при одностороннем использовании, на несколько миллионов лет. Однако растения часто дают низкие урожаи именно из-за недостатка азота в почве. Это объясняется тем, что только небольшая группа азотистых соединений может быть быстро усвоена растениями. Не только свободный азот, но и многие формы связанного азота не могут служить источником азотного питания для растений. Азот, поступающий в виде белковых веществ в почву вместе с остатками растений и животных, совсем не годится для этих целей, он должен быть подвергнут минерализации, а образующийся при этом аммиак должен быть окислен в соли азотистой и азотной кислот. В основе процессов круговорота азота лежат следующие биохимические процессы: гниение белков, разложение мочевины, нитрификация, денитрификация и фиксация атмосферного азота. (8, 159)
Гниение, или аммонификация белков — микробиологический процесс, при котором под воздействием гнилостных микроорганизмов происходит гидролитическое расщепление белков, поступающих в почву с трупами животных и отмирающими растениями, с образованием промежуточных продуктов (альбумоз, пептонов, амино- и амидокислот), а также дурно пахнущих веществ — индола, сероводорода, меркаптана, летучих жирных кислот.
Конечным продуктом гидролиза белков и дезаминирования аминокислот является NH3, почему этот процесс и называется аммонификацией белка. Таким образом, при гниении происходит минерализация белковых веществ, которая в зависимости от химического состава белков субстрата, вида гнилостных бактерий и условий их жизнедеятельности может быть полной или не доведенной до конца. При полной минерализации белка образуются СО2, NH3, Н2О, H2S и минеральные соли. При широком доступе кислорода продукты гидролиза белков подвергаются полному окислению, зловонных веществ образуется значительно меньше, чем при анаэробных условиях. Такой процесс называется тлением.
Гниение — преимущественно анаэробный процесс, при котором полного окисления некоторых продуктов, например жирных кислот, не происходит. Гнилостные микробы широко распространены в почве, воде, воздухе, в животных и растительных организмах. Поэтому любой продукт, не защищенный от них, быстро подвергается гниению. Его вызывают как анаэробные, так и аэробные микроорганизмы, причем они могут действовать и преемственно, и одновременно. Наиболее энергичными возбудителями гниения, сопровождающегося глубоким распадом белка и образованием азотистых и безазотистых соединений (индола, скатола, жирных кислот и др.), являются Bacillus mycoides, B.Mesentericus, а также Clostridium putrificum, C.sporogenes. Последние два — анаэробы, содержатся в кишечнике и после смерти вызывают зловонное разложение трупов.
Процессы гниения протекают только при наличии условий, благоприятных для жизнедеятельности их возбудителей (влажность, температура и т. п.). В сухой песчаной почве трупы подвергаются мумификации (высушиванию без гниения). Гнилостные процессы происходят и в организме человека, в частности в кишечнике; причиной их являются Е.со1i и другие микробы. По мнению И. И. Мечникова, продукты гниения (скатол, индол и др.), постоянно образующиеся в организме, вызывают хроническую интоксикацию и являются одной из причин преждевременного старения.
Гнилостные процессы протекают также при газовой гангрене: ткани, омертвевшие под влиянием образуемых возбудителями этой болезни экзотоксинов, заселяются гнилостными аэробными и анаэробными бактериями и подвергаются распаду. Некоторые гнилостные процессы используются в промышленности с полезной целью, например при выработке кожи для отделения от нее шерсти — швицевании.
Исключительное значение процессов гниения заключается в том, что они играют важную роль в естественном самоочищении почвы и воды. Этим пользуются для строительства специальных очистных сооружений (полей ассенизации, орошения и т. п.), для биологической переработки и обезвреживания фекальных нечистот и сточных вод, содержащих много мертвых белковых субстратов. Гниение ведет к обогащению почвы азотистыми продуктами. (13, 250)
Следующим важным этапом круговорота азота, вслед за образованием NH3, является процесс нитрификации, т. е. окисление NH3 вначале в азотистую, а затем в азотную кислоту, соли которых наиболее пригодны для азотного питания растений. Процесс нитрификации вызывается двумя группами открытых С. Н. Виноградским нитрифицирующих бактерий. Нитрозобактерии окисляют NH3 до азотистой кислоты, а нитробактерии окисляют азотистую кислоту в азотную.
Нитрифицирующие бактерии — строгие аэробы, хемолитотрофы. Энергию окисления они используют для восстановления СО2 в гексозу. Благодаря нитрифицирующим бактериям в почве могут образовываться огромные скопления солей азотной кислоты в виде селитры (в Чили, Перу). Завершая процесс минерализации белковых веществ, нитрифицирующие бактерии играют исключительно важную роль и в процессах самоочищения почвы и воды, и в санитарно-гигиенических устройствах (поля орошения и т. п.). Таким образом, нитрифицирующие бактерии способствуют повышению урожайности почвы благодаря накоплению в ней азотнокислых солей.
Однако в почве происходят и противоположные процессы, т.е. денитрификации, или восстановлений микроорганизмами солей азотной кислоты в соли азотистой кислоты и в другие простые азотистые соединения, вплоть до свободного азота, который, уходит в атмосферу.
Способностью восстанавливать нитраты в нитриты обладает большое количество видов бактерий и грибов.
Денитрифицирующие бактерии (в частности, некоторые виды Pseudomonas) в анаэробных условиях используют денитрификацию как основную форму дыхания. Для них соли азотной и азотистой кислот служат источниками азота. Энергию для своей специфической деятельности денитрифицирующие бактерии получают из органических веществ, которыми богата почва. Денитрифицирующие бактерии наносят вред сельскому хозяйству, так как способствуют обеднению почвы минеральным азотом и переходу свободного азота в атмосферу. Особенно энергично процессы денитрификации развиваются в слежавшейся, плохо аэрируемой почве. Однако убыль азота из почвы, вызванная активностью денитрифицирующих бактерий, компенсируется деятельностью свободноживущих аэробных и анаэробных и клубеньковых азотфиксирующих бактерий. Более 90% азота связывают азотфиксирующие бактерии: на каждый гектар почвы ежегодно от 25 до 300 кг азота привносят только они.
Так, при самом активном участии многих видов микроорганизмов, в природе происходит непрерывный круговорот азота, поддерживающий существование жизни на Земле. (16, 90)
Схема 1 «Круговорот азота».
Процессы распада безазотистых органических веществ обусловлены по преимуществу жизнедеятельностью микроорганизмов, а процессы созидательные — фотосинтезом зеленых растений, водорослей и фотосинтезирующих бактерий. В основе процессов распада безазотистых органических веществ лежат различные формы брожения, которые постоянно происходят в природе. Брожение – анаэробное дыхание, при котором микроорганизмы используют выделяющуюся энергию для своей жизнедеятельности.
Впервые биологическую природу брожения открыл в 60-х годах 19 в. гениальный французский ученый Луи Пастер. Пастеру удалось на примере молочнокислого, спиртового и маслянокислого брожения доказать, что эти процессы вызываются жизнедеятельностью микроорганизмов. (6, 81)
Спиртовое брожение углеводов вызывают дрожжи (Saccharomyces cerevisiae), некоторые виды бактерий (Sarcina ventriculi) и отдельные представители мукоровых грибов рода Mucor.При спиртовом брожении молекула гексозы распадается на этанол и углекислый газ. В ходе брожения образуется много промежуточных продуктов — гексозомонофосфат, фруктозодифосфат, фосфотриозы, фосфоглицериновая кислота, фосфопировиноградная кислота, пировиноградная кислота, уксусный альдегид и, наконец, этиловый спирт.
При содержании в сбраживаемом растворе более чем 30% сахара часть его остается неиспользованной, так как при этих условиях образуется до 15% спирта, а при такой концентрации спирт подавляет жизнедеятельность дрожжей. Поэтому натуральные вина содержат не более 15% спирта. Главное преимущество чистых культур дрожжей заключается в том, что брожение виноградного сока протекает и заканчивается быстро, а отсутствие посторонней микрофлоры позволяет получать вина хорошего вкуса и аромата (с хорошим «букетом»). По окончании брожения молодое вино стабилизируют и дают ему созреть. Эти процессы занимают несколько месяцев, а при изготовлении высококачественных красных вин — даже несколько лет. В течение первого года во многих красных винах происходит второе, спонтанное брожение — яблочно-молочнокислое, которое вызывается рядом молочнокислых бактерий (Prdiococcus, Leuconostoc). В результате этого яблочная кислота винограда превращается в молочную кислоту и СО2, т. е. дикарбоновая кислота превращается в монокарбоновую, и кислотность вина уменьшается, оно становится высококачественным.
Уксуснокислое брожение — биологический окислительный процесс, при котором с помощью уксуснокислых бактерий спирт окисляется в уксусную кислоту. Если какую-либо жидкость, содержащую небольшое количество спирта (вино, пиво), оставить открытой, то в ней постепенно появляется уксусная кислота и кожистая пленка (уксусная матка) на поверхности. Уксуснокислые бактерии объединены в род Acetobacter, содержащий ряд видов и подвидов. Этиловый спирт под влиянием уксуснокислых бактерий подвергается окислению, в результате которого вначале образуется уксусный альдегид, а затем — уксусная кислота. При использовании специальных рас уксуснокислых бактерий максимальный выход уксуса достигает 14,5%. Уксуснокислые бактерии превращают ряд многоатомных спиртов в сахар. Одна из таких реакций используется для получения сорбозы из сорбитола. Сорбоза — промежуточный продукт синтеза аскорбиновой кислоты. Она применяется в качестве суспендирующего агента при изготовлении многих лекарственных препаратов. Уксуснокислые бактерии могут наносить вред в виноделии и пивоваренной промышленности, вызывая прокисание вина и пива.
Молочнокислое брожение — широко распространенное биохимическое явление, давно известное на примере скисания молока. Под влиянием молочнокислых бактерий (семейство Lactobacillaceae) лактоза расщепляется на составляющие ее гексозы — глюкозу и галактозу, которые затем специфическими ферментами превращаются в молочную кислоту. Свертывание молока происходит вследствие того, что молочная кислота отщепляет кальций от казеина, белок превращается в параказеин и выпадает в осадок. Молочнокислые бактерии широко распространены в природе. Они обнаруживаются в молоке, воздухе, на коже, шерсти, в тонком и толстом кишечнике и представлены большим количеством видов палочковидных и кокковидных бактерий, различающихся не только по морфологии, но и физиологическим свойствам (по использованию различных источников углерода и азота).
Маслянокислое брожение также широко встречается в природе. Возбудитель маслянокислого брожения был открыт Л. Пастером. На примере маслянокислого брожения Л. Пастер разработал учение об анаэробах. Типичный представитель бактерий маслянокислого брожения — азотфиксирующий Clostridium pasteurianum. Маслянокислые бактерии в больших количествах встречаются в почве, навозе, на растениях, в молоке, сыре. Многие из них являются анаэробами и относятся к роду Clostridium.
Маслянокислое брожение — сложный биохимический процесс расщепления углеводов, в ряде случаев жиров и белков, на масляную кислоту, углекислоту и воду, при этом образуется много побочных продуктов — уксусная, молочная, пропионовая и другие кислоты.
Из числа других форм брожения чрезвычайно важным является брожение целлюлозы (клетчатки), в которой заложены огромные запасы углерода. Разложение целлюлозы, которая в количественном отношении представляет один из основных компонентов растительных тканей, осуществляется главным образом высоко специализированными в отношении питания аэробными и анаэробными микроорганизмами. Среди аэробных бактерий, расщепляющих целлюлозу, наиболее важны скользящие бактерии рода Cytjphaga. Целлюлоза — единственное вещество, которое они могут использовать в качестве источника углерода. Цитофаги быстро растворяют и окисляют целлюлозу.
Сера — составная часть некоторых белков. Одним из конечных продуктов гниения белков является H2S. Сероводород не усваивается высшими растениями. Биохимические превращения серы восстановительного и окислительного порядка осуществляются серобактериями. Для них H2S является источником энергии. Серобактерии окисляют H2S с выделением свободной серы, которая отлагается у них в цитоплазме в виде капель.
В клетках бактерий сера окисляется далее до серной кислоты, образующиеся сульфаты служат прекрасным питательным веществом для высших растений. H2Sв серную кислоту окисляют различные виды пурпурных серобактерий.
Наряду с такими сульфурирующими бактериями в природе не менее широко распространены и десульфурирующие микробы (аналоги денитрифицирующих бактерий), они восстановливают сульфаты, вызывая образование H2S. Выделение H2S десульфурирующими бактериями происходит в глубинах морей, поэтому в Черном море на глубине 2500 м содержание H2S доходит до 6,5 мл в 1 л воды. Значительное накопление H2S в результате биологического восстановления серы наблюдается в целебных грязях, в лиманах и других водоемах. В санитарном отношении серобактерии являются важными агентами начальной стадии биологического очищения сточных вод и разложения органических отбросов, содержащих серу. (2, 14)
С химической стороны круговорот фосфора достаточно прост, поскольку он встречается в живых организмах только в пятивалентном состоянии в виде свободных фосфатных ионов (РО4-3) или в составе органических фосфатных компонентов клетки. Бактерии не способны поглощать большинство органических фосфорсодержащих соединений, свои потребности в фосфоре они удовлетворяют путем поглощения фосфатных ионов, из которых затем синтезируют органические фосфатные соединения. При разложении гнилостными бактериями белковых веществ одновременно с минерализацией азота происходит превращение органического фосфора в фосфатные ионы. Поскольку большая часть фосфатов, несмотря на быстрый круговорот фосфора, находится в виде нерастворимых солей кальция, железа или алюминия, фосфаты также служат фактором, ограничивающим рост растений. Растворимые фосфаты постоянно переносятся из почвы в море вследствие выщелачивания. Этот перенос имеет однонаправленный характер. Лишь небольшая часть фосфатов возвращается на сушу, главным образом в виде отложений гуано морскими птицами. Поэтому доступность фосфатов для растений зависит от непрерывного перевода в раствор нерастворимых фосфатных отложений — процесса, в котором важную роль играют микроорганизмы. Образуемые ими кислые продукты метаболизма (органические кислоты, а также азотная и серная) растворяют фосфат кальция, а образуемый ими H2S способствует растворению фосфата железа. (3,





