

Божьи коровки (coleoptera, coccinellidae) фауны города гомеля. дипломная (вкр). биология. 2021-06-13
Министерство
образования Республики Беларусь
Учреждение
образования
«Гомельский
государственный университет имени Франциска Скорины»
Биологический
факультет
Кафедра
зоологии и охраны природы
Дипломная
работа
БОЖЬИ
КОРОВКИ (COLEOPTERA, COCCINELLIDAE) ФАУНЫ ГОРОДА ГОМЕЛЯ
Исполнитель:
Столярова Наталья Николаевна
Гомель 2021
Содержание
Введение
. Обзор литературы
.1 История изучения божьих коровок
.2 Характеристика объекта
исследования
. Объект, программа и методика
исследования
2.1 Объект исследования
2.2 Место и методы проведения
исследования
. Результаты исследований и их
обсуждения
.1 Аннотированный список и анализ
таксономической структуры
.2 Видовой состав и экологические
особенности божьих коровок исследованных территорий
.2.1 Стационар «Заливной луг»
.2.2 Стационар «Суходольный луг»
.2.3 Стационар «Опушка леса»
.3 Биотопическая приуроченность
кокцинеллид в исследованных территориях
.4 Гигропреферендум божьих коровок в
исследованных территориях
.4.1 Стационар «Опушка леса»
.4.2 Стационар «Суходольный луг»
.4.3 Стационар «Заливной луг»
Заключение
Список использованных источников
Введение
Большая часть населения в промышленно развитых
странах сосредоточена в городах. В то же время урбанизация – это процесс, полностью
изменяющий окружающую среду, так как город характеризуется значительной
мозаичностью среды, пространства, на которых проходит активная биологическая
жизнь, отделяются пространствами биологически «неактивными» и последние
являются естественными барьерами. Зеленые насаждения города разной обильности,
формы и происхождения окаймляют сеть улиц и зданий таким образом, что «живые
анклавы» среды образуют своеобразные острова, целиком изолированные друг от
друга. Они различаются происхождением и степенью воздействия антропогенного
фактора [3]. Постройка зданий, прокладка улиц, коммуникаций, обильные
промышленные эмиссии приводят к уничтожению природной среды – почвенного и
растительного покрова, животного населения. Различные антропические факторы
нарушают водный, воздушный, световой режимы, в связи с чем городские экосистемы
выделяют в разряд отдельных – так называемых урбоэкосистем [6]. Одна из главных
особенностей городской экосистемы в том, что в ней встречаются как условия,
характерные для естественных местообитаний, так и особенные, к которым
относятся: более теплый и сухой микроклимат, обширная кормовая база, а также
немалое количество «новых» свободных экологических ниш для заселения.
Отсутствие конкуренции и давления со стороны хищников и паразитов ведет к
нестабильным отношениям между видами, проникающими в городскую экосистему и
видами, ее уже заселившими.
Жесткокрылые являются важными компонентами
трофических и топических взаимодействий, многие из них имеют серьёзное
практическое значение, как вредители запасов или зелёных насаждений. Почвенная
мезафауна имеет большое индикационное значение, к примеру, она является
показателем, прежде всего таких свойств почвы, как влажность.
Жесткокрылые распространены почти по всему свету
и встречаются в самых разнообразных местообитаниях, в том числе под камнями и
бревнами, в лесной подстилке, в гравии по берегам рек и в пресных водоемах.
Божьи коровки приносят довольно много пользы в
сельском и лесном хозяйстве, так как они поедают тлю, приносящую вред зеленым насаждениям.
В связи с этим изучение видового состава, особенностей биологии и экологии
божьих коровок для более рационального применения в борьбе с вредителями весьма
актуально и привносит значительный вклад в экономическую безопасность страны. В
то же время ряд видов кокцинеллид поедают зеленые части растений, принося
определенный вред, но это не идет ни в какое сравнение с приносимой ими
пользой. Такое двойственное поведение привлекает внимание и является объектом
исследования многих исследователей.
Исследования, посвященные божьим коровкам
естественных биоценозов, а также проблемам, касающихся биологических методов
борьбы с вредителями сельского и лесного хозяйства проходят довольно широко. В
то же время изучение состояния сообществ кокцинеллид в антропогенно
трансформированных биоценозах городов – урбоценозах, незначительны и требуют
самого пристального рассмотрения.
В связи с этим целью данной работы было изучение
видового состава и структуры сообществ божьих коровок фауны г. Гомеля,
обитающих в ряде открытых биоценозов.
В ходе данной работы были поставлены следующие
задачи:
. Выявление и уточнение видового состава божьих
коровок, обитающих в урбоценозах г. Гомеля.
. Определение параметров биоразнообразия
сообществ кокцинеллид фауны г. Гомеля.
. Оценка видовой структуры и экологических
особенностей сообществ божьих коровок.
Объектом исследования являлись божьи коровки –
жесткокрылые насекомые из семейства Coccinellidae.
В качестве предмета исследования выступали
видовой состав и экологическая структура сообществ кокцинеллид.
Изучение божьих коровок носят большое
теоретическое и практическое значение, так как они являются полезными
насекомыми и регулируют численность тли в естественных экосистемах и
сообществах, созданных руками человека.
Практическое значение работы заключается в том,
что данная работа начинает собой цикл исследований, посвященных изучению
населения божьих коровок, обитающих на территории города Гомеля. Комплексное
изучение позволит выяснить экологические параметры сообщества, обитающего в
сильн6о трансформированных биоценозах городской среды. Результаты исследований
были использованы при выполнении тем ГБ 06-15 «Структура и функционирование
зооценозов наземных и водных экосистем Белорусского Полесья».
Таким образом, изучение божьих коровок носят
большое теоретическое и практическое значение.
Глава 1. Обзор литературы
.1 История изучения божьих коровок Беларуси
божья коровка кокцинеллид
гигропреферендум
Изучение божьих коровок начинается с 1902 года.
В это время Н.М. Арнольд создает «Каталог насекомых Могилевской губернии» [1].
Далее, в 1954 году Н.П. Дядечко изучил фауну кокцинеллид Украины [13].
Несколько позже Г.И. Савойская в 1983 году изучила систематику, а также
применение божьих коровок в борьбе с вредителями сельского хозяйства. Кроме
того, этот же автор сделала акцент на изучении личинок кокцинеллид фауны
бывшего СССР. Большое число представителей группы вошло в опубликованную
фундаментальную работу [27].
Изучение видового состава и биологии
непосредственно кокцинеллид Беларуси занимался на рубеже 60-ых ХХ века годов
В.П. Семьянов, обративший пристальное внимание на коровок трибы Scymnini
[31], для которых характерна, как правило, невысокая численность, локальное
распространение и низкая встречаемость. В 1968 году В.П. Семьянов изучил
биологию Calvia decemguttata [30] а в 1969 году – население, биологию и
полезную роль кокцинеллид фауны Беларуси [33]. В 1980 году он продолжил
изучение биологии Calvia decemguttata [32] а в 1981 рассмотрел новые данные о
паразитах и хищниках Coccinella
septempunctata [29].
Спустя три года (в 1984 году) он обратил внимание на изучение методики учета
кокцинеллид, их паразитов, патогенов и хищников [34], а также в октябре этого
года В.П. Семьянов опубликовал свою статью «Кокцинеллиды агроценозов Белорусии»
[35].
Значительный вклад в изучение божьих коровок
внес также В.И. Курилов [18]. Он дал оценку эффективности кокцинеллид в
снижении численности тлей на картофеле в условиях нашей страны. В 1983 году
С.В. Буга и Е.С. Шалапенок изучали фауну и стациальное распределение
кокцинеллид заказника «Налибокская пуща», а в 1991 году они же опубликовали
комплексную работу в виде обширной сводки по фауне представителей этого
семейства на территории нашей страны – «Обзор фауны кокцинеллид (Coleoptera,
Coccinellidae) Белоруссии» [9,
10].
Таким образом, фауна кокцинеллид изучалась,
прежде всего, с точки зрения защиты сельскохозяйственных культур от вредителей.
Чисто фаунистических работ (за исключением сводки С.В. Буги и Е.С. Шалапенок)
для территории Беларуси практически нет, что позволило нам начать изучение этой
хозяйственно важной группы жесткокрылых.
.2 Характеристика объекта исследования
Кокцинеллиды (Coccinellidae)
– одно из крупнейших семейств отряда желтокрылых (Coleoptera),
насчитывающий более 5000 видов, из которых около 2000 встречаются в
Палеарктике. На территории бывшего СССР отмечен 221 вид. В Беларуси встречается
56 видов. Божьи коровки распространены во всех частях света. Одни из них
встречаются на всех растениях: деревьях, кустарниках или травах, на которых
только есть тли; другие держатся только на полевых травах; третьи – на лугах,
прилегающих к ручьям; четвёртые – только на деревьях; наконец, некоторые виды
живут на тростнике и на других водяных растениях; последние отличаются более
длинными ногами, которые помогают им держаться на растениях, легко гнущихся от
ветра.
Жуки небольших размеров – длина тела имаго от 1
до 18 мм. Тело обычно округло-овальное, сильно выпуклое (рисунок 1), почти
полушаровидное (нижняя сторона почти плоская или слабовыпуклая). У некоторых
групп тело продолговато-овальное, в той или иной степени уплощенное.
Поверхность чаще всего голая, реже – покрыта волосками.

Рисунок 1 – Внешний вид Coccinella
septempunctata
Систематическое положение семиточечной божьей
коровки:
Царство Zoa
Подцарство Metazoa
Тип Arthropoda
Подтип Tracheata
Надкласс Hexapoda
Класс Insekta-Ectognata
Отряд Coleoptera
Подотряд Polyphaga
Надсемейство Lamellicornia
Семейство Coccinellidae
Голова небольшая, может быть вытянута в
продольном или поперечном направлении. Глаза большие, часто с выемкой на
переднем крае. Усики 8-11 члениковые, короткие или средней длины, с булавой
(чаще) или без нее. Переднее- и среднегрудь поперечные. Заднегрудь широкая,
почти квадратная, гораздо длиннее среднегруди. Ноги умеренной длины, покрыты
густыми волосками. Лапки скрыто 4-члениковые (кажутся 3-члениковыми, так как
3-й членик маленький и скрыт в лопастях 2-го) и только у представителей трибы Lithophilini
лапки явственно 4-члениковые. Переднеспинка шире головы, выпуклая, поперечная,
с вырезкой разной формы на переднем крае. Часто- с пятнами или рисунком из
слившихся пятен. Надкрылья красные, желтые, коричневатые с черными или белыми
пятнами, которые, иногда сливаются, образуют изменчивый рисунок; или надкрылья
черные с красными или желтыми пятнами. Брюшко снизу почти совсем плоское,
сверху гораздо более плоское, чем надкрылья, и состоит из 5-6 видимых
стернитов. Половой диморфизм выражен слабо. У большинства видов вершина 5-го
или 6-го стернита у самцов с вырезкой или ямкой, у самок – с бугорком. У
некоторых видов узор отличаются по рисунку на переднеспинке [2].
Божьи коровки очень плодовиты и в зависимости от
местных условий и особенностей вида откладывают от 40 до 700 и более яиц
небольшими группами по 10-30 штук на листья или другие части растений.
Яйца обычно овальные, слегка сужены к концу. У
видов триб Stethorini
и Chilocorini – короткие, почти
округлые. Окраска яиц желтая, оранжевая, беловатая; поверхность часто
шагренирована. Яйцекладки обычно плотные, яйца расположены более или менее
правильными рядами, касаясь друг друга боками. У некоторых особей Hamonia
sedecimnotata яйцекладки
«рыхлые», яйца отодвинуты друг от друга на расстояние, равное 1-1.5 диаметра
яйца (пока это единственный известный мне случай такого типа яйцекладки) [4].
Личиночный период божьих коровок длится три
недели, в течение которого каждая личинка истребляет несколько сотен тлей.
Личинки более или менее камподеовидные, удлиненные, иногда плоские и овальные.
У личинок коровок, питающихся червецами, тело покрыто восковидными нитями
белого цвета. Личинки часто пестро окрашены, рисунок образуют часто оранжевые,
желтые или белые пятна. Поверхность тела покрыта волосками, щетинками,
бородавками и другими выростами. Личинки в своем развитии проходят 4 возраста
[5].
Куколки свободные, прикрепляются к субстрату
остатками экзувия личинки. Часто имеют яркую окраску с черными и белыми
пятнами. Для трибы Coccinellini
характерен открытый тип – куколка расположена в лопнувшей со спинной стороны
личиночной шкурке. У Chilocorini
полузакрытый тип – личиночные покровы лопаются частично и обнажают лишь спинку
куколки. У Hyperaspini
куколки находятся под личиночной шкуркой [7].
На нижней стороне листа самка откладывает свои
оранжевые яички кучками до полусотни в каждой. Всего одна самка способна
отложить до 600 яиц. Через одну-две недели из них выходят черные с желтыми
пятнами личинки размером 2-3 мм. Через месяц, достигнув в длину сантиметра, тут
же в зоне питания личинка семиточечной божьей коровки превращается в черную
неподвижную куколку. А еще через неделю-полторы из треснувшей вдоль спины
шкурки из нее выходит имаго.
Осенью божьи коровки перелетают с полей и лугов
на опушки леса, к заросшим травой берегам рек и оврагам. Здесь, под корой, во
мху, под опавшими листьями они проводят зиму. [8].
Большинство из божьих коровок – активные
хищники. Божьи коровки уничтожают массу различных вредителей сада и поля –
тлей, червецов, щитовок, медяниц, мелких личинок и гусениц. Например, за период
своего развития личинка семиточечной божьей коровки съедает до 800 тлей, а
взрослый жук – до 50. Другие виды божьих коровок съедают также огромное
количество вредителей. Питающиеся тлей божьи коровки активны весной и летом,
особенно в теплые солнечные дни, они находятся в непрерывном поиске своей
добычи.
В то же время сами божьи коровки несъедобны для
большинства насекомоядных птиц и других животных из-за специфического запаха и
едкой гемолимфы. В случае опасности жуки поджимают сяжки под голову, а ноги под
туловище, притворяются мёртвыми и выпускают желтоватый острый сок, прежде
употреблявшийся против зубной боли. Виды, имеющие более длинные ноги, в таких
случаях стараются спастись бегством.
Божьим коровкам, несмотря на их название,
свойственна склонность к каннибализму. Особенно она проявляется у личинок. Но и
взрослые особи нередко поедая яйца своего вида. Яйца съедаются даже отложившей
их самкой, если она не находит другой пищи. Нехватка пищи свидетельствует о
том, что ее может не найтись и для личинок, которые вылупятся из яиц и без
корма скоро погибнут. Съедая яйца, самка пополняет свои ресурсы, необходимые
для откладки последующих порций яиц, в более удобном месте или в более
подходящее время. У голодающей самки возможна задержка яйцекладки, и она не
сможет воспользоваться более благоприятной для ее потенциального потомства
ситуацией [15].
Для зимовки божьи коровки подыскивают места
среди густой растительности, листового опаду, под корой сухих деревьев и т.д.
или внутри помещений, сараев, навесов. Они часто залетают в дома, гнездятся
между дверьми, двойными оконными рамами, в складках штор.
Высокая эффективность и быстрое нарастание
численности божьих коровок при благоприятных условиях делают их одним из
естественных факторов биологической борьбы с вредными насекомыми. В целях
сохранения и повышения активности божьих коровок применение пестицидов должно
строго регламентироваться, а в период активности коровок и других полезных
насекомых целесообразно воздерживаться от химических обработок и применять
биологические средства защиты растений, которые снижают численность вредных
насекомых, не оказывая отрицательного действия на полезную энтомофауну. Это
позволит естественным врагам восстанавливать свои популяции и контролировать
численность вредителей. [14].
Божьи коровки выступают классическими объектами
популяционных исследований на основе фенетических и эволюционно-экологических
подходов. Этому способствуют их широкое распространение, массовая встречаемость
и характерный полиморфизм по окраске и рисунку покровов.
В трофическом отношении у кокцинеллид выделяются
следующие группы [12]:
афидофаги (питаются тлями),
кокцидофаги (питаются червецами и щитовками),
акарифаги (питаются клещами),
фитофаги (питаются растительной пищей).
В свою очередь фитофаги подразделяются на:
филлофагов, которые питаются листьями, реже
цветками или плодами;
палинофагов, питающихся пыльцой растений;
мицетофагов, питающихся мицелием грибков.
В семействе Cocinellidae
обычно выделяются 7 подсемейств [12]:
Sticholotidinae
(= Sticholotinae)
Coccidulinae
Tetrabrachinae
(= Lithophilinae) – иногда
включается в Coccidulinae
Scymninae
Chilocorunae
Coccinellinae
Epilachninae
Наиболее массовым представителем семейства можно
считать семиточечную божью коровку Coccinella
septempunctata (см.
рисунок 1).
2. Объект, программа и методика исследований
.1 Объект исследований
Объектом исследования являлись божьи коровки,
обитающие на трех стационарах – «Опушка леса», «Суходольный луг» и «Заливной
луг».
Божьи коровки как представители животного мира
имеют следующее систематическое положение:
Царство Zoa
Подцарство Metazoa
Тип Arthropoda
Подтип Tracheata
Надкласс Hexapoda
Класс Insecta-Ectognata
Отряд Coleoptera
Подотряд Polyphaga
Семейство Coccinellidae
[16].
2.2 Место и методы проведения исследования
Исследования проводились стационарно с мая по
сентябрь 2009 – 2021 года на трёх участках, которые находились в районе
микрорайона «Солнечный». Стационары представляли собой:
1 «Опушка леса». Стационар наименее подверженный
рекреационной нагрузке. Расположен на границе города. Растительность
представлена мятликовым разнотравьем, из травянистых растений преобладают
одуванчик лекарственный – Taraxacum
officinale
Wigg., клевер пашенный
– Trifolium
arvense L.,
клевер ползучий – T. repens
L. Подрост
составляли молодые сосны и берёзы. Почва- супесь тяжёлая.

Рисунок 2 – Стационар «Опушка леса»
«Заливной луг». Стационар покрыт большим
количеством прибрежной растительности. Рекреационная нагрузка незначительная,
преимущественно встречаются рыбаки. Растительность представлена снытью
обыкновенной – Aegophodium
padagrara
L., лютиком жгучим- Ranunculus
flammula
L. и лапчаткой
серебристой- Potentila
argenthea
L. Почва- суглинок
лёгкий.

Рисунок 3 – Стационар «Заливной луг»
3 «Суходольный луг». Характеризуется
значительной рекреационной нагрузкой, т.к. расположен недалеко от проезжей
части. Из растительность преобладают мятликовые. Почва- супесь легкая.

Рисунок 4 – Стационар «Суходольный луг»
Сбор материала проводился при помощи кошения
сачком. Энтомологический сачок (рисунок 8) состоял из трёх частей: обруч, сетка
и рукоятка. Обруч изготавливался из стальной проволоки сечением 4 мм. У
стандартного сачка для кошения диаметр обруча 30 см. Сетка, или мешок был сшит
из мельничного газа (капроновой сетчатой ткани).
В качестве рукоятки сачка использовалась
деревянная палка длиной 1,5 м.

Рисунок 5 – Сачок [17].
Собранные, при помощи кошения сачком, насекомые
помещались в морилку для умерщвления. Морилка представляла собой специальное
устройство для умерщвления насекомых. Современные морилки изготавливаются из
стеклянных или пластиковых стойких к растворителю широкогорлых банок с плотной крышкой.
Объём произвольный и зависит от размера и количества помещаемых в морилку
насекомых. На дне располагается либо ватный тампон, зашитый в ткань, либо кусок
поролона таким образом, чтобы он не вываливался при переворачивании банки.
Заполняли объём морилки куском белой х/б или льняной ткани, который прекрасно
впитывает избыток влаги, препятствует повреждению насекомых и не нуждается в
частой замене. Морилка заправляется ядом. С прошлого века пользуются обычным
медицинским эфиром или хлороформом. Насекомые после замаривания этими ядами
получаются жёсткими и почти не поддаются расправлению и монтировке для
коллекции (рисунок 6).
Рисунок 6 – Морилка
Лучше всего использовать эфиры уксусной кислоты
– этиловый, амиловый, бутиловый. Каждая морилка была этикетирована из расчета 1
учет – 1 морилка.
На каждом из стационаров проводилось по 4 учета,
которые представляли собой по 25 двойных взмахов сачком. Собранные божьи
коровки в лаборатории выкладывались из морилки на ватные слои (матрасики) для
дальнейшего хранения и определения(рисунок 7). Идентификация видов жесткокрылых
проводилась с использованием определительных таблиц [19].
Рисунок 7 – Матрасик
Для изготовления матрасика использовался
прямоугольный кусок ваты небольшой толщины, который помещался в бумагу. Внутрь
вкладывалась этикетка, на которой подписывался стационар, на котором
производился сбор материала, число сбора, и обнаруженные виды. Насекомые
выкладывались в четыре ряда, в соответствии с методикой сбора, каждый ряд
соответствовал одному прокосу.
Для статистической обработки количественных
показателей использовались пакеты прикладных программ. Первичная база
жесткокрылых составлялась с использованием «MS Excel». Для анализа
распределений, средних, ошибок и верификации гипотез об их различиях и связях
использовался пакет «Statistica 6.0». Показатели α-разнообразия
в сообществах [43, 44, 45, 46, 47] были рассчитаны с использованием
программного пакета «BioDiversity Pro». Расчет индекса разнообразия Шеннона
проводился с использованием натурального основания логарифма.
Доминирование в сообществе определялось по шкале
Ренконена [35], согласно которой виды, относительное обилие от общего числа
особей которых составляет более 5%, считаются доминантными; 2-5% –
субдоминантными; 1-2% – рецедентными; менее 1% – субрецедентными.
Типология ареалов определялась нами исходя из
номенклатуры типов распространения, предложенной Городковым [11].
Материал хранится на кафедре зоологии и охраны природы
УО «Гомельский государственный университет имени Франциска Скорины».
3. Результаты исследований и их обсуждения
.1 Аннотированный список и анализ
таксономической структуры
На основе собственных и литературных данных был
составлен аннотированный список для 38 видов божьих коровок, обитающих в
открытых урбоценозах г. Гомеля, который включал в себя следующие харатеристики:
название, распространенные синонимы, тип ареала, биотопическую приуроченность,
зимующую фазу, срок генерации [20, 21, 22].
СЕМЕЙСТВО COCCINELLIDAE
LATREILLE, 1807
Триба Madaini
Gordon, 1975
Подсемейство Epilachninae
Триба Epilachnini
Род Subcoccinella
Agassiz, 1846
S. vigintiquatuorpunctata
(Linnaeus, 1758).
Палеарктический вид. Луговой мезофил: луга переходного типа и суходолы, рудеральная
растительность, агроценозы. Тяготеет к ксерофитным стациям, обычный, местами
(на юго-западе республики) фоновый вид. Фитофаг, имаго и личинки повреждают
маревые, некоторые бобовые и особенно растения родов Silene
L. и Melandrium
Roehl (Caryophillaceace)
[13]. Может вредить люцерне и высадкам свеклы. 1-2 генерации в год. Перед
зимовкой жуки образуют скопления.
Подсемейство Coccidulinae
Mulsant, 1846
Триба Coccidulini
Род Coccidula
Kugelann, 1798
C. rufa
(Herbst, 1783).
Палеарктический вид. Пойменные и переходного типа луга, посевы зерновых и
кормовых злаков, низинные болота, гигрофил. Вполне обычный, местами фоновый
вид. Гербофил, хищник преимущественно поабионтных тлей на растениях семейств Poaceae
и Cyperaceae. Зимует имаго под
корой деревьев, в дуплах и других убежищах, т.е. местах зимовок климатического
типа [14], часто вдали от типичных стаций обитания.
C. scutellata
(Herbst, 1783).
Палеарктический вид. Гербобионт. Пойменные луга, болото, гигрофильный вид.
Встречается локально, преимущественно около водоемов, немногочислен. Афидофаг,
хищник тлей на вейниках, маннике, троснике, осоках. Имаго зимуют под корой
деревьев, в дуплах, нередко с C.
rufa.
Подсемейство Scymninae
Mulsant, 1846
Триба Scymnini
Mulsant, 1846
Род Scymnus
Kugelann, 1794
S. (Pullus) ater Kugellan, 1794. Европейский
вид. Лиственные леса, лесной мезофил. Редкий, малочисленный, локально
встречающийся вид; более характерен для юга Белорусии. Дендробионт. Хищник
дендрофильных тлей, главным образом на дубах.
S. (Pullus)
auritus Thunberg,
1795. Западнопалеарктический вид. Смешанные и широколиственные леса, парки.
Дендробионт. Считается степным мезофиллом [15]. В условиях региона типичный
консорт дуба черешчатого, распространение которого предопределяет
распространение коровки: местами это обычный и весьма многочисленный вид.
Афидофаг, личинки специализированы к питанию тлями сем. Phylloxeredae.
В светлых дубравах жуки изредка образуют предзимовочные скопления, но зимовка
имаго группами не отмечалась.
6 S. (Pullus) terrugatus (Moll,
1785). Транспалеаркт.
Лиственные
леса, лесной мезофилл. Редок. Хищник преимущественно филлобионтных тлей сем. Drepanosiphidae
(Callaphididae) и подтрибы Rhopalosiphina
(Aphididae)
S. (Pullus)
suturalis
Thunberg, 1795. Распространен
по всей Палеарктике. Хвойные леса. Дендробионтный вид, тесно связанный с
сосной. Вполне обычный, как правило, немногочисленный в Белоруссии. Считается
степным мезофиллом (Мизер, 1974). Афидофаг.
8 S. (s. str.) nigrinus Kugellan,
1794. Европейско-сибирский
вид. Хвойные и смешанные леса, лесной мезофил. Встречается по всей территории
республики. Обычный, но не массовый вид. Дендробионт, связан с тлями сем. Lachnidae
(Schizolachnus
pineti (F.),
Cinara spp.)
на соснах. Жуки Sc.
Nigrinus нередко (в отличии
т большинства сцимнин) присуствуют в послезимовочных скоплениях других видов
кокцинеллид [16].
9 S. (s. str.) rubromaculatus
(Goeze, 1777). Рспространение: Европейско-Сибирская
область и область Древнего Средиземноморья (ОДС). Смешанные и широколиственные
леса, перелески, сады, парки, лесной мезофил. Более характерен для юга
республики. Дендробионт. Хищник дендрофильных тлей сем. Drepanosiphidae
и Aphididae [17].
S. (Nephus)
bipunctatus
Kugellan, 1794.
Европейско-сибирский вид. Лиственные леса. Относительно редок. Хищник кокцид
[18].
Триба Stethorini
Род Stethopys
Weise 1885
S.punctillum
Weise, 1861.
Распространен по всей Палеарктике. Смешанные и лиственные леса, сады, зеленые
насаждения – повсюду, где присуствуют клещи-фитофаги сем. Tetranychidae,
которыми питаются имагои личинки коровки. Лесной мезофил, предпочитающий
ксеротермные условия. Вид обычен для всей территории Белоруссии, но на высоком
уровне численности отмечается лишь местами и в отдельные годы.
Подсемейство Chilocorinae
Mulsant, 1846
Триба Platynaspini
Род Platynaspis
Redtenbacher, 1846
P. luteorubra
(Goeze, 1777).
Южнопалеарктический вид. Лиственные и смешанные леса, лесной мезофил. Довольно
редок. Дендробтонт. Хищник кокцид.
Триба Chilocorini
Mulsant, 1846
Род Chilocorus
Leach, 1815
C. renipustulatus
(Scriba, 1790).
Транспалеарктический вид лесной мезофил встречается повсюду, где присутствуют
дендрофильные диаспиновые щитовки, фоновый вид. Высокая численность отмечается
в очагах щитовок сем. Diaspididae
(Chionaspis
salicis L.,
Diaspidiotus
alni March.,
Lepidosaphes
ulmi L.,
и др.). личинки обычны в сборах кошением по кустарничкам [19]. Скоплений не
образует, но жуки концентрируются на участках коры с высокой плотностью жертв.
C.bipustulatus
(Linnaeus, 1758).
Распространен по всей Палеарктике. Лесной мезофил. Повсюду встречается вместе с
Ch. Renipustulatus,
значительно уступаю ему по уровню численности. Различия в параметрах экониш
этих видов хилорин в условиях региона трудноуловимы. По-видимому, несколько
более толеранен к низкой плотности жертв и чаще встречается вне выраженных
очагах диаспиновых щитовок [37].
E.quadripustulatus
(Linnaeus, 1758).
Европейско-сибирский вид. Леса, перелески, сады, зеленый насаждения и т.п.,
лесной мезофил. Дендрофильный, практическиэвритопный представитель группы.
Встречается повсюду, но численность редко бывает значительной. Характеризуется
широким кругом трофических связей, включающим представителей Coccidoidea
и Aphidoidea, примечательна
тенденция к питанию ложнощитовками (Coccidae)
и их экологическими аналогами среди дендрофильных тлей сем. Hormaphididae
[20].
Подсемейство Coccinellinae
Latreille, 1807
Триба Coccinellini
Род Coccinula
Dobzhansry, 1925
H. tredecimpunctata
(Linaeus, 1758).
Голарктический вид. Характерен для травянистой растительности и кустарников.
Гигрофил. Распространен повсеместно, фоновый вид. Хищник тлей на злаках,
осоках, зонтичных и многих околоводных растениях [38]. Встречается и на
надводной растительности.
A. novemdecimpunctata
(Linnaeus, 1758).
Голарктический вид. По биотопической приуроченности нет существенных различий
между данным видом и двумя предыдущими, в то же время четче прослеживается
связь 19-точечной коровки с поафильными тлями сем. Aphididae.
Распространен повсеместно, фоновый вид.
A. variegata
(Goeze, 1777).
Распространен в Палеарктике, проникает в Индо-Малайскую область. Термофильный
вид, встречающийся на суходольных лугах, участках рудеральной растительности,
изреженных посевах с.-х. культур, повсеместно, но многочислен лишь местами.
Численность популяций в сильной степени зависит от погодных условий, вследствие
чего наблюдаются значительные колебания ее уровня по годам. Хищник
преимущественно ксерофильных тлей трибы Macrosiphini
(Aphididae) [21]. Жуки зимуют
под куртинками полыни, подстилке на сухих возвышенных местах, иногда небольшими
группами.
S. undecimnotata
(Schneider, 1792). Выходец из
области Древнего Средиземноморья. Проникает по азональным стациям в лесную
зону. Суходольные луга, другие стации с ксеротермными условиями, степной
мезофилл [39]. Очень редок. Хищник ксерофильных тлей. Для фауны Белоруссии
указывается впервые.
A. bipunctata
(Linnaeus, 1758).
Гоарктический вид. В условиях региона – один из наиболее массовых
представителей семейства. Виду свойственна широкая экологическая валентность, в
то же время ето дендрофил (лесной мезофил). Встречается во всех стациях при
условии произрастания древесных растений и присуствия жертв (дендрофильные
тли). Численное доминирование A.
bipunctata в комплексах
кокцениллид (по показателю относительного обилия) достоверно свидетельствует о
том, что данный комплекс претерпел антропогенную структурную трансформацию (Honek,
Rejmanek, 1982). Как
высокоантропотолерантная форма является преобладающим видомкоровок в стациях,
находящихся под сильным антропогенным воздействием (городские зеленые
насаждения и т.п.) [41, 42]. Вид полиморфен: особи, тносящиеся к f.
typica, состовляют в
популяциях от 70 до 90% [22]. Доля особей, принадлежащих к основным “черным”
формам (f. quadrimaculata,
f. sexpustulata),
может достигать 30%. Другие меланисты (f.
lunigera, f.
sublunata), а также
многоточечная форма (f.
annulata) представлены
единичными экземплярами. В Белоруссии, как и в других европейских регионах, не
выявлена f. impunctata.
A. decempunctata
(Linnaeus, 1758).
Палеарктический вид. Леса, сады, зеленые насаждения и т.п., лесной мезофил.
Обычен повсюду, но сравнительно не многочислен. Тесно связан с дендрофильными
тлями (Drepanosiphidae,
Chaitophoridae, Aphididae)
лиственных пород. Для имаго весьма характерна зимовка в неопавших скрученных
листьях дуба черешчатого (позднераспускающаяся форма) [40]. Вид полиморфный,
красные формы встречаются несколько чаще желтых (черных).
C. septempunctata
Linnaeus, 1758.
Распространение: Палеарктическая и Индо-Малайская области. Массовый эвритопный
вид. Тяготеет к травянистой растительности. Основной объект питания – тли
разных систематических и экологических групп, очень хорошо (как у личинок, так
и имаго) выражена склонность к миксоэнтомофагии [23]. Жуки могут образовывать
пред- и послезимовочные скопления.
C. quinquepunctata
Linnaeus, 1758.
Палеарктический вид. Перелески, луга, поля, вырубки, обочины дорог и т.п.,
луговой мезофил. Фоновый для всей территории Белоруссии вид. Тяготеет к
травянистой растительности. Хищник негаллообразующих тлей. Жуки часто образуют
скопления.
C. saucerotti
lutshniki
dobrzhansky, 1917. Подвид
имеет сильно фрагментированный ареал, по-видимому, в пределах Европейского
континента. Номинативный подвид распространен в Восточной Азии. В Восточной
Европе коровка Лучника, очевидно, имеет ареал ленточного типа (по долинам рек).
Единственное известное местонахождение в Белоруссии – пойма р. Сивичанки (на
территории заказника «Налибокская пуща»). Коровка тесно связана с ивами,
произрастающими в речных поймах. Хищник ивовых тлей. Для фауны Белоруссии
указывается впервые.
C. magnifica
Redtenbacher, 1849.
Палеарктический вид. Разреженные сосновые леса, суходольные луга, степной
мезофил. Редок, более характерен для юго-запада региона. Гербофильный и
мирмекофильный вид, встречается практически только около муравейников.
C. hieroglyphica
Linnaeus, 1758. Широко
распространен в Палеарктике. Форма, типичная для верховых и переходных болот. В
Белоруссии вид имеет мозаичное распространение, но его, по-видимому, можно
отнести к числу редких лишь на юге республики. Психрофильный гигрофил. Круг
объектов питания включает личинок листоедов подсемейства Galerucinae,
а также тлей на кустарниках, злаках и осоках. Места зимовки жуков –
климатологического типа.
C. quatuordecimpustulata
(Linnaeus, 1758).
Транспалеарктическй вид. Суходольные и переходного типа луга, поля, луговой
мезофил. Гербебионт. В условиях региона в ксеротермных стациях является
массовой формой. Хищник поафильных тлей.
O. conglobata
(Linnaeus, 1758).
Голарктический вид. Смешанные и лиственные леса, перелески, вырубки, сады,
зеленые насаждения и т.п., лесной мезофил. Вслед за A.
bipunctata проникает в
уличные зеленые насаждения городов. Обычен, но редко достигает высокого уровня
численности. Дендробионт. Хищник тлей (Drepanosiphidae,
Aphididae) на лиственных
деревьях. Весьма примечательны частые находки жуков в неопавших листьях дуба
(нередко вместе с A. 10-punctata),
т.е. в местах зимовок гипсотактичского типа.
H. quadripunctata
(Pontoppidan, 1837).
Западнопалеарктический вид. Хвойные и смешанные леса лесной мезофилл.
Встречается местами и немногочислен. Дендрофильный вид, связанный с тлями сем. Lachnidae
и Adelgidae, преимущественно
на соснах.
M. octodecimguttata
(Linaeus, 1758).
Европейско-Сибирский вид, не выходит за пределы лесной зоны. Хвойные и смешанные
леса, лесной мезофилл. В Белоруссии обычен повсюду. Дендрофильный вид, тесно
связанный с тлями сем. Adelgidae
и Lachnidae на елях. Жуки
зимуют в пнях, дуплах, под корой деревьев, образуя массовые скопления, часто
совместно с другими коровками данной трибы (особенно A.
obliterate).
S. vigintiguttata
(Linnaeus, 1758).
Европейский вид. Широколиственные и смешанные леса, лесной мезофилл.
Встречается местами, несколько чаще – в южных и юго-западных районах
республики. Дендробионт. Связан с дендрофильными тлями сем. Drepanosiphidae
и некоторыми листоблошками на лиственных деревьях. Для него, по-видимому,
характерен весьма невысокий уровень антропотолерантности.
C. quatuordecimguttata
(Linnaeus, 1758). Широко
распространен в Голарктике. Лиственные и смешанные леса, перелески, сады,
зеленые насаждения, лесной мезофилл. Фоновый вид. Дендробионт.
Специализированный хищник листоблошек. Период ухода жуков на зимовку сильно
растянут (вторая половина июля- октябрь). Отдельные экземпляры иногда
присутствуют в образуемых другими видами кокцениллид зимовочных скоплениях.
C. decemguttata
(Linnaeus, 1767).
Транспалеарктический вид. Широколиственные и смешанные леса, перелески, сады,
зеленые насаждения, лесной мезофил. Считается неморальной формой. В Белоруссии
относительно редок. Чаще встречается на дубе черешчатом и лещине. Хищник
дендрофиьлных тлей (Drepanosiphidae)
и листоблошек.
C. quinquedecimpunctata
(Fabricius, 1777).
Распрастранен в Палеарктике, проникает в Индо-Малайскую область. Лиственные
леса, сады, лесной мезофилл. Редок. Дендробионтный вид. Kруг
объектов питания очерчен не очень четко, считается, что в него входят
дендрофильные тли (Drepanosiphisae),
псиллиды и, возможно, личинки листоедов [36].
P. quatuordecimpunctata
(Linnaeus, 1758).
Распространен в Палеарктике и Эфиопской области. Массовый эвритопный вид,
численно доминирующий (среди кокценеллид) в травяном ярусе практически всех
стаций, кроме ксеротермных. Хищник гербофильных тлей. Скоплений не образует.
A. ocellata
(Linnaeus, 1758).
Голарктический вид. Леса, перелески, зеленые насаждения, сады, лесной мезофил.
Повсюду обычен. Дендрофил, связанный с тлями преимущественно на хвойных, реже –
лиственных деревьях [24].
Триба Psylloborini
Род Psylabora
Linnaeus, 1758.
P. vigintiduopunctata
(Linnaeus, 1758). Транспалеарктический
вид. Леса, вырубки, перелески, зеленые насаждения, сады и т.д. Эвритопный вид.
Обычен повсюду, местами массовая форма. Личинки- облигатные микофаги, связанные
с аскомицетами сем. Erysiphaceae.
Это также основной объект питания имаго [26]. Образование предзимовочных
скоплений весьма характерно для азиатской части ареала. В условиях региона это
довольно редкое явление, которое можно наблюдать на хорошо прогреваемых
участках разреженных лиственных лесов, лесных полянах. Жуки зимуют в подстилке
либо группами в сухих трухлявых пнях, изредка- под корой деревьев [28].
38 H.
sedecimguttata (Linnaeus,
1758). Транспалеарктический вид. Смешанные и широколиственные леса, зеленые
насаждения. Лесной мезофилл. Вид не принадлежит к числу редких, но встречается
спорадически и всегда немногочислен, особенно на севере Белоруссии [25]. Это
единственный представитель семейства, у которого имаго активно летят на свет.
3.2 Видовой состав и экологические особенности
божьих коровок исследованных территорий
За время исследований на трех стационарах нами
было отловлено 651 экземпляр божьих коровок. Наибольшей численностью
представителей семейства Coccinellidae
(247) характеризовался стационар «Суходольный луг» (таблица). Несколько ниже
численность божьих коровок была отмечена на стационаре «Заливной луг» – 209
особей (таблица), а наименьшее количество экземпляров было характерно для
стационара «Опушка леса» – всего 195 особей божьих коровок (таблица).
Практически подобный характер имело также
распределение по видовому богатству. Так, наибольшее число видов было
зафиксировано на стационаре «Суходольный луг» – 17 видов (таблица). Одинаковое
число видов (по 12) было отмечено на стационарах «Заливной луг» и «Опушка
леса». Таким образом, исходя из полученных нами результатов, выявлено, что как
по видовому богатству, так и по численности божьи коровки преобладали на
стационаре «Суходольный луг». Это в определенной степени можно объяснить тем,
что основная масса встреченных видов – мезофилы, в связи с чем переувлажненные условия
заливного луга не являются оптимальными для обитания большинства собранных
видов коккцинеллид. Опушка леса, кроме того, что переувлажнена в отличие от
суходольного луга, так к тому же, еще затенена, что в некоторой степени
препятствует нормальному развитию кормовой базы тлей – основного пищевого
объекта божьих коровок как зоофагов. Необходимо отметить, что в результате
исследований в урбоценозах города Гомеля нами были обнаружены виды, ранее в
литературе не указанные для Гомельского региона [9]. Это Halyzia
sedecimguttata, отмеченная на стационарах «Заливной луг» и «Суходольный луг», а
также Hippodamia notata, которая была зафиксирована также на двух стационарах –
«Суходольный луг» и «Опушка леса».
Таблица 1 – Видовой состав и степень
доминирования (%) кокцинеллид исследованных территорий
Вид | Стационары | ||
Заливной | Суходольный | Опушка | |
Adalia | 2,87 | 2,43 | 4,62 |
Adalia | 0 | 0,4 | 0 |
Calvia | 1,91 | 1,21 | 0,51 |
Calvia | 0,96 | 1,21 | 0 |
Calvia | 0 | 0,4 | 0 |
Exochomus | 0 | 0,4 | 0 |
Coccinella | 6,22 | 5,67 | 3,59 |
Coccinella | 24,4 | 39,61 | 32,82 |
Coccinula | 19,62 | 6,48 | 15,9 |
Halyzia | 0,48 | 0,4 | 0 |
Hippodamia | 0 | 2,02 | 1,54 |
Hippodamia | 19,14 | 2,93 | 9,74 |
Myrrha | 0 | 0,4 | 0 |
Oenopia | 0 | 0 | 0,51 |
Propylaea | 11,48 | 22,27 | 22,57 |
Psyllobora | 0,48 | 3,24 | 1,54 |
Subcocinella | 1,44 | 1,62 | 4,1 |
Tyttaspis | 11 | 9,31 | 2,56 |
Всего | 209 | 247 | 195 |
Всего | 12 | 17 | 12 |
Информационное | 0,86 | 0,84 | 0,82 |
Информационное | 0,16 | 0,22 | 0,2 |
Выравненность | 0,35 | 0,3 | 0,33 |
Видовая структура божьих коровок на всех трех
исследованных стационарах варьирует в значительной степени. Так,
видами-доминантами, которые преобладали на всех трех изученных участках были
Coccinella septempunctata, Propylaea quatuordecimpunctata и Coccinula
quatuordecimpustulata. При этом численность их была хоть и высокой, но не
равнозначной для всех стационаров. Первый из рассматриваемых видов (Coccinella
septempunctata) уменьшал свое относительное обилие по мере увеличения влажности
на биотопах – от 39,61 до 24,4% на суходольном и заливном лугу соответственно
(таблица). Второй общий доминант (Coccinula quatuordecimpustulata) наоборот, имел
наименьшее относительное обилие на стационаре «Суходольный луг» – всего 6,48%.
На опушке леса ее численность возросла вдвое и составила уже 15,9%. На самом
влажном из исследованных участков (стационар «Заливной луг») ее численность
была самой высокой (в три раза выше, чем на суходольном лугу!) – 19,62%.
Численность Propylaea quatuordecimpunctata также варьировала, но судя по
параметрам, приведенным таблице, распространение этого вида в меньшей степени
зависит от влажности (относительное обилие на суходольном лугу и на опушке леса
практически идентичны – 22,27 и 22,57% соответственно)
При анализе степени доминирования божьих коровок
на стационаре «Заливной луг», кроме выше упомянутых видов доминировал также и
Hippodamia tredecimpunctata. Численность этого вида в сравнении с другими
доминировавшими видами было невелико и было на уровне численности Coccinula
quatuordecimpustulata (относительное обилие – 19,14%)
На стационаре «Суходольный луг», кроме
упомянутых выше Coccinella septempunctata, Propylaea quatuordecimpunctata и
Coccinula quatuordecimpustulata более доминантов
не наблюдалось. Кроме того, Hippodamia tredecimpunctata,
доминировавший на стационарах «Заливной луг» и «Опушка леса» на рассматриваемом
участке имел довольно низкую численность (относительное обилие – 2,93%) – в
четыре раза меньшую чем на опушке леса и в восемь (!) раз на заливном лугу, что
может свидетельствовать о предпочтении этим видом более влажных местообитаний.
Также следует отметить, что на данном стационаре было отмечено больше всего
видов, которые более нигде из исследованных территорий не были встречены. Это
такие виды как Adalia decimpunctata, Calvia quinquedecimpunctata, Exochomus
quadripustulatus и Myrrha octodecimguttata (рисунок 6). Относительное обилие
этих видов было невелико (0,4%), но, в то же время, они вносят свою долю в
более высокое видовое разнообразие стационара. По этому показателю можно
косвенно определить более благоприятные условия обитания для божьих коровок в
Гомеле.

Рисунок 8 – Внешний вид божьих коровок: А –
Adalia decimpunctata, Б – Exochomus quadripustulatus, В – Myrrha
octodecimguttata
На стационаре «Опушка леса» среди преобладающих
видов наблюдались 4: Coccinella septempunctata, Coccinula
quatuordecimpustulata, Hippodamia tredecimpunctata и Propylaea
quatuordecimpunctata. При этом за исключение последнего упомянутого вида, все
доинирующие виды имели промежуточную численность в сравнении с другими
стационарами (таблица).

Рисунок 9 – Внешний вид Oenopia conglobata
Как особенность условий обитания на этом
стационаре можно отметить отсутствие на нем таких видов как Calvia
quatuordecimguttata и Halyzia sedecimguttata, которые, хоть и в небольшом
количестве, но были встречены на двух других исследованных территориях. В то же
время на данном участке был встречен вид Oenopia conglobata (рисунок 7),
который на других стационарах не фиксировался.
.2.1 Стационар «Заливной луг»
Видовое разнообразие на стационаре «Заливной
луг» в 2009 и 2021 годах практически не различаются. Если в 2009 году
количество видов, обнаруженных на данном стационаре составило всего 9, то уже в
2021 среди обнаруженных божьих коровок было выявлено 10 видов. Проанализировав
видовое разнообразие кокцинеллид за два года можно увидеть, что общее
количество составило 12 видов (таблица 2
Таблица 2 – Видовой состав и степень
доминирования (%) кокцинеллид
Вид | Год | |
2009 | 2021 | |
Adalia | 3,85 | 2,29 |
Calvia | 5,13 | 0 |
Calvia | 0 | 1,53 |
Coccinella | 2,56 | 8,4 |
Coccinella | 29,49 | 21,37 |
Coccinula | 21,79 | 18,32 |
Halyzia | 1,28 | 0 |
Hippodamia | 32,06 | 11,45 |
Propylaea | 0 | 18,32 |
Psyllobora | 0,76 | |
Subcocinella | 1,28 | 1,53 |
Tyttaspis | 2,56 | 16,03 |
Всего | 9 | 10 |
Всего | 78 | 131 |
Если сравнивать процентное соотношение видов
божьих коровок, обнаруженных в 2009/2021 годах на стационаре «Заливной луг» то
можно увидеть что оно варьирует в той или иной степени. Такие виды как Calvia
quatuordecimguttata, Propylaea quatuordecimpunctata, Psyllobora
vigintiduopunctata на данном стационаре не встречались в 2009 году, но, в
относительно небольшом количестве, были обнаружены в 2021 году. И наоборот виды
Halyzia sedecimguttata и Calvia decemguttata были встречены в 2009 году, а в
2021 – не обнаружены. Остальные виды божьих коровок, указанные в таблице
встречались два года подряд на данном стационаре. В 2009 году такие виды как Adalia
bipunctata, Coccinella
septempunctata, Coccinula
quatuordecimpustulata
и Hippodamia tredecimpunctata преобладали в процентном соотношении, и являлись
доминантными на данном стационаре. В 2021 году к этим доминантам прибавились
еще два вида Propylaea quatuordecimpunctata и Coccinella quinquepunctata.
.2.2 Стационар «Суходольный луг»
Видовое разнообразие на стационаре «Суходольный
луг» было наиболее богатым и составило 17 видов. Как и на предыдущем стационаре
было различие в количестве видов в 2009 и 2021 году. На первый год исследования
было обнаружено всего 12 видов, но уже в 2021 году видовое разнообразие
достигло цифры 15.
Таблица 3 – Видовой состав и степень
доминирования (%) кокцинеллид на стационаре «Суходольный луг» за весь период
исследований
Вид | Год | ||
2009 | 2021 | ||
Adalia | 3,57 | 1,84 | |
Adalia | 0 | 0,61 | |
Calvia | 1,19 | 1,23 | |
Calvia | 0 | 1,84 | |
Calvia | 0 | 0,61 | |
Exochomus | 0 | 0,61 | |
Coccinella | 8,33 | 4,29 | |
Coccinella | 47,62 | 35,58 | |
Coccinula | 5,95 | 6,75 | |
Halyzia | 1,19 | 0 | |
Hippodamia | 2,38 | 1,84 | |
Hippodamia | 1,19 | 3,68 | |
Myrrha | 1,19 | 0 | |
Propylaea | 25,01 | 20,88 | |
Psyllobora | 1,19 | 4,29 | |
Subcocinella | 1,19 | 1,84 | |
Tyttaspis | 0 | 14,11 | |
Всего | 12 | 15 | |
Всего | 84 | 163 | |
Что же касается относительного обилия, то в 2009
году почти половину всех особей составил вид Coccinella septempunctata
(47,62%), в 2021 году оно было незначительно ниже и составило – 35,58%. К числу
доминантов как в 2009, так и в 2021 году следует отнести такие виды как
Propylaea quatuordecimpunctata, Coccinella quinquepunctata и Coccinula
quatuordecimpustulata. Их относительное обилие в 2009 году составляло 25,01%,
8,33% и 5,95%, и 20,88%, 4,29% и 6,75% в 2021 году соответственно.
Также в 2021 году к числу доминантов можно
отнести вид, который не встречался в 2009 – Tyttaspis sedecimpunctata, его
относительное обилие составило 14,11%. Также, помимо доминанта (Tyttaspis
sedecimpunctata) в 2021 году были обнаружены следующие виды не встреченные на
первый год исследования – Adalia decimpunctata, Calvia quatuordecimguttata,
Calvia quinquedecimpunctata, Exochomus quadripustulatus. В 2009 году также
присутствовали виды, которые исчезли с данного стационара в 2021 году. Это виды
– Myrrha octodecimguttata, Halyzia sedecimguttata.
.2.3 Стационар «Опушка леса»
Количество видов, обнаруженных на данном
стационаре за два года исследования составило, как и на стационаре «Заливной
луг» – 12. В 2009 году из восьми встреченных видов два вида в 2021 году не
встречены, это такие виды как Calvia decemguttata и Oenopia conglobata. В 2021
году было обнаружено четыре вида которых не было в 2009 году, это в первую
очередь доминант 2021 года Propylaea quatuordecimpunctata (33,59%), Psyllobora
vigintiduopunctata, Subcocinella vigintiquatuorpunctata и Tyttaspis sedecimpunctata.
Таблица 4 – Видовой состав и степень
доминирования (%) кокцинеллид на стационаре «Опушка леса» за весь период
исследований
Вид | Год | |
2009 | 2021 | |
Adalia | 6,25 | 3,82 |
Calvia | 1,56 | 0 |
Coccinella | 7,81 | 1,53 |
Coccinella | 43,75 | 27,46 |
Coccinula | 12,5 | 17,56 |
Hippodamia | 1,56 | 1,53 |
Hippodamia | 25,01 | 2,29 |
Oenopia | 1,56 | 0 |
Propylaea | 0 | 33,59 |
Psyllobora | 0 | 2,29 |
Subcocinella | 0 | 6,11 |
Tyttaspis | 0 | 3,82 |
Всего | 8 | 10 |
Всего | 64 | 131 |
Если сравнивать результаты исследований на всех
трех стационарах, то можно обнаружить небольшую закономерность, что в 2021
году, на всех трех стационарах наблюдалось значительное увеличение как числа
видов, так и числа особей. Это можно объяснить более выгодными погодными
условиями, которые, по сравнению с 2009 годом, способствовали лучшему развитию
как кормовой базы, так и мест их обитания.
Для более полного анализа биоразнообразия божьих
коровок нами было проведено ранжирование обнаруженных видов кокцинеллид. При
анализе графика «ранг-обилие» (рисунок 8) можно сказать, что распределение
видов по типу «разломанного стержня» характерно только для одного стационара –
«Суходольный луг». Лог-нормальное распределение более характерно для стационара
«Заливной луг», а стационар «Опушка леса» занимает промежуточное положение.
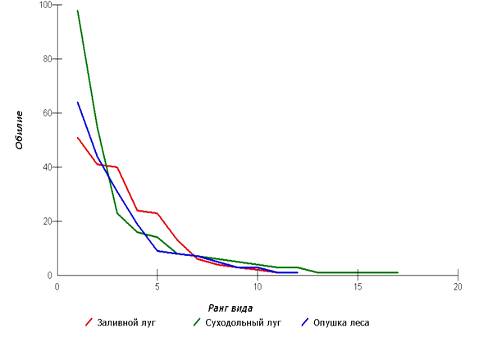
Рисунок 10 – Ранжирование видов божьих коровок
на исследованных стационарах
Выявленная особенность может говорить о том, что
наиболее оптимальные условиях обитания божьих коровок из всех исследованных
урбоценозов наблюдаются на стационаре «Суходольный луг» и они характеризуется
низкой конкуренцией и широким видовым богатством.
.3 Биотопическая приуроченность кокцинеллид
исследованных территорий
При оценке биотопической приуроченности тех или
иных видов к местам обитания (биопреферендума) был выявлен ряд особенностей.
Так, на стационаре «Суходольный луг» по видовому богатству (рисунок 9) было
определено, что преобладали лесные виды (Adalia bipunctata, Calvia
decemguttata, Hippodamia tredecimpunctata и др.). В несколько меньше количестве
были встречены собственно луговые виды (Coccinella quinquepunctata, Coccinula
quatuordecimpustulata, Tyttaspis sedecimpunctata и др.). Меньше всего было
зафиксировано эвритопных видов (Coccinella septempunctata, Propylaea
quatuordecimpunctata и др.)
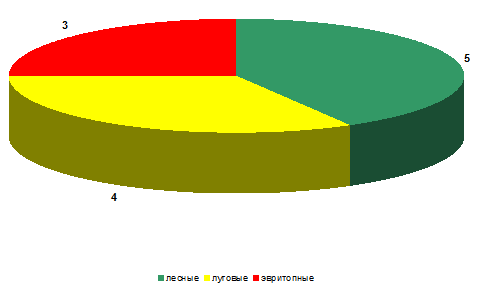
Рисунок 11 – Биопреферендум кокцинеллид на
стационаре «Заливной луг» (видовое богатство)
В то же время, несмотря на преобладающее видовое
богатство лесных видов по численности (рисунок 11) они уступают как луговым
(относительное обилие – 39%), так и эвритопным видам (относительное обилие –
36%). Таким образом, можно сказать, что на стационаре складывается равномерное
распределение всех групп кокцинеллид по биотопической приуроченности.
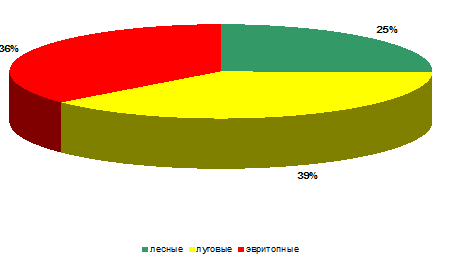
Рисунок 12 – Биопреферендум кокцинеллид на
стационаре «Заливной луг» (относительное обилие)
На стационаре «Суходольный луг» ситуация
складывается иная. Так, по видовому богатству (рисунок 12) лесные виды (Adalia
bipunctata, Calvia decemguttata, Calvia quatuordecimguttata, Hippodamia notata,
Myrrha octodecimguttata и др.) практически вдвое превышают число луговых видов
(Coccinella quinquepunctata, Coccinula quatuordecimpustulata, Tyttaspis
sedecimpunctata и др.) и в три раза – эвритопных (Coccinella septempunctata, Propylaea
quatuordecimpunctata и Psyllobora vigintiduopunctata).
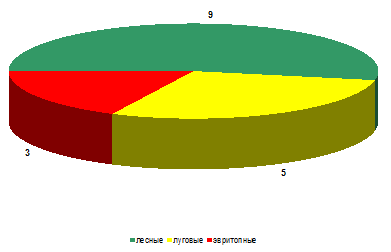
Рисунок 13 – Биопреферендум божьих коровок на
стационаре
«Суходольный луг» (видовое богатство) При
оценке численности наблюдается совершенно иные особенности (рисунок 13).
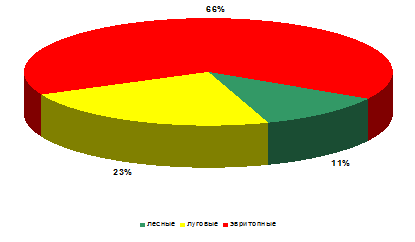
Рисунок 14 – Биопреферендум божьих коровок на
стационаре «Суходольный луг» (относительное обилие)
Относительное обилие лесных видов кокцинеллид на
суходольном лугу крайне резко отличается от их видового богатства и значительно
уступает (в 6 раз) эвритопным видам.
Численность луговых видов также велика в
сравнении с лесными видами (относительное обилие – 23%), но в три раза уступают
эвритопам.
Таким образом, можно сказать, что условия
обитания стационара «Суходольный луг» наиболее благоприятны для луговых и, в
первую очередь, эвритопных видов.
На стационаре «Опушка леса» показатели
биотопической приуроченности среди видового богатства (рисунок 14) идентичны с
показателями стационара «Заливной луг», что косвенно может говорить о том, что
условия увлажнения сходны на этих участках.
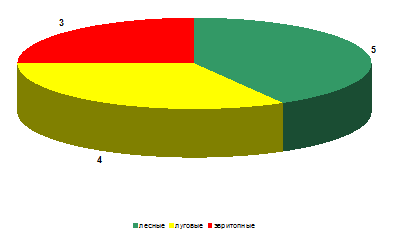
Рисунок 15 – Биопреферендум божьих коровок на
стационаре «Опушка леса» (видовое богатство)
На рассматриваемом стационаре по численности (рисунок
15), как и на двух ранее проанализированных стационарах преобладали эвритопные
виды (относительное обилие составило 57% от всех обнаруженных особей божьих
коровок).
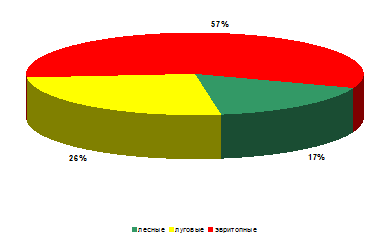
Рисунок 16 – Биопреферендум божьих коровок на
стационаре «Опушка леса» (относительное обилие)
Высока также была и численность луговых видов
(относительное обилие – 26%). Относительное обилие собственно лесных видов несколько
уступает луговым и составило 17%. Это можно объяснить экотонными особенностями
местообитания и значительным присутствием в подобных экосистемах как собственно
лесных, так и луговых видов.
Таким образом, можно сказать, что по показателю
биотопической приуроченности наиболее сбалансировано отражены именно луговые
виды (Coccinula quatuordecimpustulata, Subcocinella vigintiquatuorpunctata,
Tyttaspis sedecimpunctata, Exochomus quadripustulatus и др.), которые имеют
практически равный характер, как по видовому богатству, так и по численности на
всех изученных стационарах. Значительная численность эвритопных видов при
незначительном видовом богатстве может говорить о высокой способности данных
немногочисленных видов приспосабливаться к самым различным условиям обитания
исследованных стационаров. Низкую численность при относительно высоком видовом
богатстве лесных видов можно. По всей вероятности, объяснить узкой
специализацией к среде обитания, связанную с повышенной влажностью и
затененностью, характерную для лесов.
.4 Гигропреферендум божьих коровок в
исследованных сообществах
.4.1 Стационар «Опушка леса»
На опушке леса большую роль по видовому составу
играли мезофильные виды – 11 (рисунок 17).
В гораздо меньшей степени представлены гигрофилы
(их всего 1 вид – Hippodamia tredecimpunctata). Мы можем предложить, что для
данного стационара гигрофилы являлись нехарактерными видами.
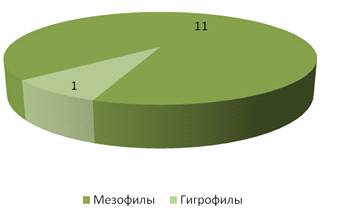
Рисунок 17 – Гигропреферендум кокцинеллид на
стационаре «Опушка леса» (виды)
Исходя из результатов, представленных на рисунке
18 можно наблюдать, что наиболее высокое видовое богатство характерно для
мезофильных видов (11 видов из 12). Это, в первую очередь такие виды, как С.
septempunctata и др. предпочитающие места с нормальным увлажнением.
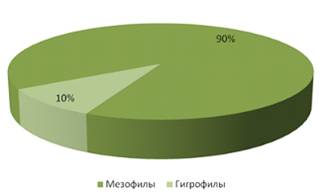
Рисунок 18 – Гигропреферендум кокцинеллид на
стационаре «Опушка леса» (особи)
На стационаре «Опушка леса», как и на
стационарах «Суходольный луг» и «Заливной луг», преобладали мезофилы – 90%, так
как мезофилы предпочитают места с нормальным увлажнением. В незначительном
количестве встречались гигрофилы – 10%, но они представлены в большем
количестве, чем на стационаре «Суходольный луг». Можно предположить, что это
связано с тем, что стационар «Опушка леса» место более увлажненное, чем
стационар «Суходольный луг».
.4.2 Стационар «Суходольный луг»
На суходольном лугу большую роль по видовому
составу играли мезофильные виды – 16 (рисунок 18). В гораздо меньшей степени
представлены гигрофилы (их всего 1 вид – Hippodamia tredecimpunctata).
Мы можем предложить, что для данного стационара
гигрофилы являются нехарактерными видами. Исходя из результатов, представленных
на рисунке 19 можно наблюдать, что наиболее широко представлены мезофильные
виды, предпочитающие места с нормальным увлажнением
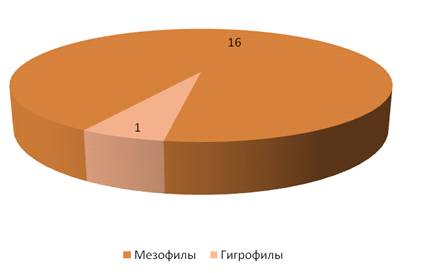
Рисунок 19 – Гигропреферендум кокцинеллид на
стационаре «Суходольный луг» (виды)
На стационаре «Суходольный луг» в большом
изобилии встречались мезофилы – 97%, что и следовало ожидать, так как
излюбленными местами мезофилов являются места с нормальным увлажнением
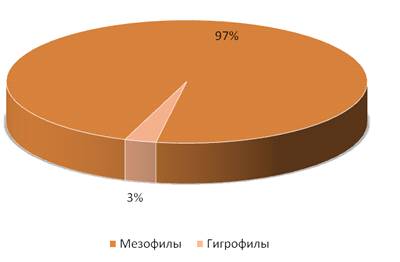
Рисунок 20 – Гигропреферендум кокцинеллид на
стационаре «Суходольный луг» (особи)
Гигрофилы, по относительному обилию, были
встречены в незначительном количестве, всего 3%, так как предпочитают места
более увлажненные.
.4.2 Стационар «Заливной луг»
На заливном лугу большую роль по видовому
составу играли мезофильные виды – 11 (рисунок 21).
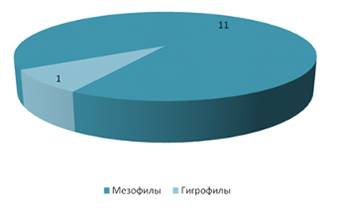
Рисунок 21 – Гигропреферендум кокцинеллид на стационаре
«Заливной луг» (виды)
В гораздо меньшей степени представлены гигрофилы
(их всего 1 вид – Hippodamia tredecimpunctata). Гигрофильный вид присутствуют
на данной территории, так как неподалёку располагалась небольшая речка.
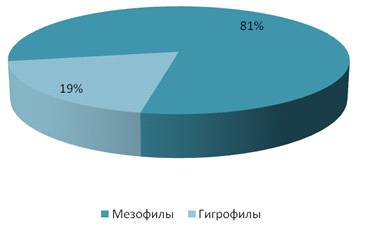
Рисунок 22 – Гигропреферендум кокцинеллид на
стационаре «Заливной луг» (особи)
Исходя из данных представленных на рисунке 22
можно наблюдать, что широко представлены мезофильные видов, их относительное
обилие составило 81%. Обилие особей гигрофильного вида составило 19%.
Причиной такого распределения особей является
то, что мезофильные виды предпочитают места с нормальным увлажнением, но не
зависимо от этого распространяются повсеместно.
В заключении можно сказать, что и по видовому
богатству и по относительному обилию преобладали виды, предпочитающие места с
нормальным увлажнением (мезофилы). Гигрофилы встречались в незначительном
количестве, так как предпочитают более увлажненные места, чем рассмотренные
стационары.
Заключение и выводы
Таким образом, в результате проведенных
исследований, можно сделать следующие выводы:
. В исследованных трех открытых биоценозах
города Гомеля зафиксировано 18 видов божьих коровок.
. Наиболее широко по видовому богатству и
численности были представлены божьи коровки на участке суходольного луга, менее
оптимальными условиями обитания оказались те, которыми характеризовалась опушка
пригородного леса. Средние показатели имел участок заливного луга поймы р. Сож
в пределах г. Гомеля.
. Среди божьих коровок преобладали по видовому
богатству лесные виды, а по численности – эвритопные. Наиболее сбалансировано
были представлены луговые виды, которые имели высокую численность и видовое
богатство на всех исследованных территориях.
В целом стационар «Суходольный луг», как
показали наши исследования, имел наиболее благоприятные условия для обитания
божьих коровок, так как имел достаточную кормовую базу, оптимальное увлажнение
и другие микроклиматические особенности для существования кокцинеллид. Это
отразилось в распределении видов и особей зафиксированных божьих коровок по
типу «разломанного стержня», что свидетельствует о низкой конкуренции.
Так как городские открытые местообитания (газоны
и разделительные зеленые полосы магистралей) по своим микроклиматическим
параметрам в целом схожи с таковыми естественных суходольных лугов, можно
высказать предположение, что трансформированные условия городских экосистем не
будут выступать препятствием для нормального существования представителей
рассмотренного семейства в Гомеле.
Список использованных источников
1. Александрович, О.Р. Каталог
жесткокрылых Белоруссии/ О.Р. Александрович, И.К. Лопаткин, А.Д. Писаненко,
В.А. Цинкевич, С.М. Снитко. – Мн: Фонд фундаментальных исследований Республики
Беларусь, 1996. – 125 с.
. Арнольд Н.М. Каталог насекомых
Могилевской губернии. / Н.М. Арнольд. – СПб., 1902
. Арнольди, Л.В. Жесткокрылые, или
жуки – Coleoptera. / Л.В. Арнольди – Животный мир СССР., т. 4. М. – Л., 1953. –
С. 425- 485.
. Бей-Биенно, Г.Я. Общая
энтомология. / Г.Я. Бей-Биенно. – М: Высшая школа, 1980. – 370 c.
. Божьи коровки / Захаров И.А.
[электронный ресурс] условия доступа:
http://zoometod.narod.ru/plav/plavil_17.html. Дата доступа: 25.02.2021. – Время
доступа: 19:30
. Божья коровка/ Ижевский С.С.
[электронный ресурс] условия доступа: http://entomolog.narod.ru/metodiki.html.
Дата доступа:01.03.2021. – Время доступа: 20:00.
. Божьи коровки / Семьянов В.П.
[электронный ресурс] условия доступа:
http://www.zin.ru/Animalia/Coccinelidae/rus/slrulist.htm. Дата доступа:
19.02.2008. – Время доступа: 18:50.
. Бондаренко, Н.В. Практикум по
общей энтомологии/ Н.В. Бондаренко, А.Ф. Глущенко. – Ленинград: Колос, 1972. –
334 с.
. Буга, С.В. Обзор фауны кокцинеллид
(Сoleopteran Coccinellidae) Белоруссии // Фауна и экология жесткокрылых Белоруссии.
/ С.В. Буга, Е.С. Шалапенок. – Мн.: Навука i тэхнiка, 1991. – С. 111-121.
. Буга, С.В. Фауна и стациальное
распределение кокцинеллид заказника «Налибокская пуща» // Биологические основы
освоения, реконструкциии и охраны животного мира Беларуси /С.В. Буга, Е.С.
Шалапенок. – Тез. докл. 5 зоол. конф. Белоруской ССР, 1983. – С. 142-143.
. Городков К.Б. Типы ареалов
насекомых тундры и лесных зон Европейской части СССР / Ареалы насекомых
Европейской части СССР. Атлас. Карты 179-221. – Л: Наука, 1984. – С.14-23.
. Горностаев, Г.Н. Насекомые СССР. /
Г.Н. Горностаев. – М: Мысль, 1970. – 353 с.
. Дядечко, Н.П. Кокцинеллиды
Украинской СССР/ Н.П. Дядечко. – Киев: Навуковая думка, 1954. – 157 с.
. Жизнь животных. Беспозвоночные/
Л.А. Зенкевич [и др.]; под редакцией Л.А. Зенкевича. – М: Просвещение, 1969. –
3Т 573 с.
. Захваткин, Ю.А. Курс общей
энтомологии / Ю.А. Захваткин. – М: Агропромиздат, 1986. – 319 с.
. Золотин, А.З. Насекомые служат
человеку. / А.З. Золотин. – Киев: Навуковая думка, 1986. – 140 с.
. Клюге, Н.Ю. Современная
систематика насекомых. /Н.Ю. Клюге. – СПб: Лань, 2000. – 336 с.
. Курилов, В.И. Оценка эффективности
кокцинеллид (Сoleopteran Coccinellidae) и других энтомофагов в снижении
численности тлей на картофеле в условиях Белорусии / В.И. Курилов –
Биологический метод борьбы с вредителями растений, 1968 – С. 225-228.
. Мамаев Б.М. Определитель насекомых
Европейской части СССР. / Б.М. Мамаев, Л.Н. Медведев, Ф.Н. Правдин. – М:
Просвещение, 1976. – 294 с.
. Мизер А.В. Обзор семейства
жуков-кокцинеллид фауны СССР. / А.В. Мизер. – Энтомол. обзор. Т. 53, вып. 3,
1974. – С.572-579.
. Насекомые / Б.В. Верещагин [и
др.]; под редакцией Б.В. Верещагина. -. М: Штиленца, 1983. – 200 с.
. Негробов, О.Н. Определитель
семейств насекомых. / О.Н. Негробов, Ю.И. Черненко. – Воронеж: Воронежский
университет, 1990. – 181 с.
. Общая и сельскохозяйственная
энтомология. / Труды всесоюзного энтомологического общества. – М: Колос, 1983.
– 270 с.
. Определитель насекомых Дальнего
Востока СССР. Т. III./ Жесткокрылые, или жуки. Ч. 1. – Л.; Наука. 1989. – 572
с.
. Радкевич А.И. Фауна, биология и
полезная роль кокцинеллид (Coleoptera, Coccinelliolae). / А.И. Радкевич. -Тр.
Витеб. пед. ин-та. Вып. 2, 1940. С. – 55-93.
. Росс, Г.Ч. Энтомология / Г Росс,
Ч.Росс, Д.Росс. – М: Мир, 1985. – 356 с.
. Савойская, Г.И. Личинки
кокцинеллид фауны СССР. / Г.И. Савойская. – Л.: Наука. 289 c.
. Савойская, Г.И. Кокцинеллиды:
Систематика, применение в борьбе с вредителями сельского хозяйства. / Г.И.
Савойская. – Алма-Ата, 1983. – 157 с.
. Семьянов, В.Н. Новые данные о
паразитах и хищниках Coccinella septempunctata L. (Coleoptera, Coccinellidae)
//Вопросы общей энтомологии. / В.Н. Семьянов. – Ленинград: Наука, 1981. – С.
11-14.
Восстановление лесной растительности на месте cуходольных лугов
УДК 581.55
ВОССТАНОВЛЕНИЕ ЛЕСНОЙ РАСТИТЕЛЬНОСТИ НА МЕСТЕ СУХОДОЛЬНЫХ ЛУГОВ
О 2021 М.Ю. Тиходеева, В.Х. Лебедева
Санкт-Петербургский государственный университет Поступила 15.03.2021
Проведен сравнительный анализ изменения биотопа и растительности суходольных лугов при зарастании их мелколиственными породами: березой пушистой (Betula pubescens), осиной (Populus trémula), ольхой серой (.Alnus incana) разного возраста; выстроен ряд увеличения эдификаторной силы древесных синузий.
Ключевые слова: структура сообществ, биотоп, эдификатор, луга, леса
Суходольные луга своим происхождением, как правило, связаны с деятельностью человека и используются как сенокосные или выпасные угодья. При прекращении или значительном сокращении их регулярной эксплуатации такие луга начинают интенсивно зарастать преимущественно мелколиственными древесными породами: березой пушистой и повислой (Betula pubescens, В. pendula), осиной (Populus trémula), ольхой серой (Alnus incana), ивой (Salix spp.). Процессы формирования древесных сообществ на месте луговых сопровождаются изменением условий биотопа (под биотопом понимается совокупность факторов среды, трансформируемых биоценозом, в частности, растительностью): преобразуется режим увлажнения, освещения, почвенного плодородия, что в конечном итоге приводит к кардинальным изменениям в составе и структуре растительного покрова. И в конечном итоге наблюдается постепенный переход некогда преобразованной человеком растительности к ее коренному типу.
Цель данной работы – сравнение характера изменения растительности суходольных лугов после полного прекращения их эксплуатации при зарастании березой пушистой, осиной и ольхой серой. Для ее достижения были поставлены следующие задачи: 1) оценить влияние березы, осины, ольхи разного возраста зарастания на изменение биотопических показателей (условий освещения, влажности, температурного режима воздуха, почвы и подстилки); 2) проанализировать интенсивность влияния этих пород на преобразование растительного покрова.
Исследования проводились в 2008-2021 гг. на суходольных лугах на территории Нижне-Свирского государственного природного заповедника. Растительность заповедника типична для Ленинградской области и представлена преимущественно лесами и болотами. Лугов мало – это суходольные луговые сообщества, возникшие в результате хозяйственной деятельности человека вблизи ранее существовавших деревень. После организации заповедника в 1980 г. и прекращения сенокошения и выпаса, луга
Тиходеева Марина Юрьевна, к.б.н., в.н.с. лаборатории геоботаники, e-mail: marinaur@list.ru; Лебедева Вера Христофо-ровна, к.б.н., в.н.с. лаборатории геоботаники, e-mail: vera_christ@mail.ru
стали постепенно де градировать, зарастая мелколиственными породами – ольхой, осиной, березой и ивой. На сегодняшний день сохранились значительные луговые массивы лишь в окрестностях Лахтин-ского залива. Эти крупнотравно-лисохвостные сообщества можно отнести к ассоциации Alopecuretum pratensis, близкой по флористическому составу к асс. Cerastio-Deschampsietum [1]. В их составе помимо лисохвоста Alopecurus pratensis (встречаемость N=0,92; покрытие максимальное/среднее (%) S=80/25), преобладает Anthriscus sylvestris (N=0,90; S=40/12), Veronica chamaedrys (N=0,91; S=60/17), Angelica sylvestris (N=0,88; S=60/13), Ranunculus ac-ris (N=0,71; S=40/10), Melampyrum nemorosum (N=0,80; S=40/14), Centaurea phrygia (N=0,62; S=70/17). Заметную роль в покрове играют злаки: Festuca pratensis (N=0,78; S=70/21) и F. rubra (N=0,86; S=45/14), Poa pratensis (N=0,87; S=35/12), Agrostis tenuis (N=0,64; S=50/12), Phleum pratense (N=0,53; S=60/13).
Для изучения изменения показателей биотопа и растительности под влиянием березы, осины и ольхи в луговых фитоценозах были выделены зоны с разным возрастом зарастания, представляющие собой суходольный луг и возрастные синузии исследуемых мелколиственных пород в 10, 20, 40 и 60 лет. При описании древостоя указывался породный состав, возраст, высота, диаметр стволов деревьев и их плотность, учитывался подрост (преимущественно березы и ели) и подлесок, в состав которого вошли: ива козья (Salix caprea), рябина (Sorbus aucuparia), черемуха (Padus avium). В каждой зоне с помощью цифрового фотоаппарата измерялась сквозистость, с использованием термогигрометра определялась температура и влажность почвы, подстилки и воздуха. На учетных площадках 0,1 м2 фиксировалось обилие и видовой состав травостоя, покрытие и толщина опада и ветоши (всего описано более 1500 учетных площадок). При обработке собранных материалов использовались методы дисперсионного анализа: значения квадратов корреляционных отношений (г)2) показали силу влияния, а коэффициент корреляции Пирсона (г) и тренд эмпирических линий регрессии определили знак связи [2].
Осина и береза благодаря способности к быстрому размножению и росту являются типичными
пионерными породами. Это деревья первой величины, отличающиеся сравнительно невысокой долговечностью. Редко можно встретить чистые древостой из этих пород старше 100 лет. Ежегодно продуцируя огромную массу семян, а осина еще и активно размножаясь корневыми отпрысками, они быстро осваивают вырубки, заброшенные сенокосные луга и пашни. Все выше сказанное относится и к ольхе серой. Но ольха серая – дерево второй величины или крупный кустарник, и еще менее долговечна (обычно возраст отдельных стволов ограничивается 60 годами). При формировании древесного яруса осины максимальная плотность (12000 шт./га) зафиксирована в 20-летней синузии. Это инвазионная стадия развития, для нее свойственно превалирование верхушечного роста. Здесь нет дифференциации деревьев, конкуренция слабо выражена. В 40-летних древостоях наблюдается разрастание крон в ширину, возрастает конкуренция, начинается разделение особей на господствующие и угнетенные, наблюдается элиминация угнетенных особей и, как следствие, снижается плотность древостоя (до 2900 шт./га). В 60-летней синузии плотность осины продолжает снижаться, при этом сохраняются только господствующие особи. Последующее разрежение (80-летняя синузия) сопровождается вхождением осины в стадию естественного отмирания, что сопровождается разрушением целостности древостоя и образованием прорывов («окон») в его пологе. Такие изменения отражаются и на условиях освещения: наименьшая сквози-
стость (освещенность) зафиксирована в 40-летней синузии (24 %), к 60 годам она возросла до 32 % и достигла максимума в 80-летней синузии (40 %). Аналогичные изменения наблюдаются в показателях древостоя (плотности и сквозистости) и в процессе разрастания березняка. В сероолыпанике, несмотря на снижение плотности с 12800 до 6000 шт./га с увеличением возраста синузий, прорывов в пологе не образуется. Кроны оставшихся деревьев разрастаются, еще плотнее смыкаясь друг с другом, и сквозистость (освещенность) последовательно падает в следующем ряду: 96 % – 45 % – 20 % – 10 %. В ходе зарастания луга березой, осиной и ольхой наблюдается тенденция к снижению температуры и увеличению влажности на всех уровнях (воздуха, подстилки, почвы) (табл. 1). Обращает внимание отклонение от общей тенденции изменение влажности подстилки и почвы в березняке: по сравнению с лугом она падает, в отличие от осинника и ольшаника. Развитие древесных сообществ определяет характер и обилие подстилки: количество опада возрастает, а ветоши – снижается. В сероолыпанике покрытие опада достигает 100 %, а толщина подстилки за счет ежегодного неполного разложения и накопления листового опада по сравнению с лугом возрастает в 3 раза, на фоне снижения количества ветоши в десятки раз. В осиннике и березняке толщина подстилки за счет быстрого разложения листового опада и сокращения обилия ветоши меньше, чем на лугу (табл. 2).
Таблица 1. Параметры биотопа луга и 60-летних синузий березы, осины и ольхи
Параметры Луг/Береза Луг/Осина Луг/Ольха
Сквозистость, % 98/31 98/30 96/10
Воздух температура, С° 31/27 24/21 27/21
влажность, % 56/62 55/67 50/76
Подстилка температура, С° 27/26 25/18 26/21
влажность, % 89/69 79/92 72/87
Почва температура, С° 24/26 22/18 26/22
влажность, % 76/73 80/93 78/86
Проективное покрытие опада, % 0/78 1/74 0/100
Проективное покрытие ветоши, % 83/23 85/8 84/0
Примечание: в таблице представлены средние значения
В 40-летнем осиннике начинает формироваться подлесок из рябины и черемухи, появляется подрост березы. По мере старения и разрежения древесного яруса роль подлеска постоянно возрастает (от 1800 до 5200 шт./га), при этом меняется обилие и состав подроста: от 60-летнего к 80-летнему осиннику в 2 раза возрастает участие березы и развивается 25-30-летний подрост ели с плотностью 200 шт./га. Наличие устойчивого подроста ели свидетельствует о начальных этапах развития коренного елового леса. Аналогичные результаты получены и при анализе развития подлеска и подроста в березняке, с той лишь разницей, что подрост ели появился раньше и в 60-летнем березняке его плотность уже достигла 400 шт./га. В подлеске олыпа-
из 25 измерений
ника отмечены ива козья, рябина и черемуха. Плотность подлеска в 60-летнем ольшанике для экземпляров высотой от 3 м составила 4400 шт./га. Подрост ели и ее всходы отсутствовали. Следует также отметить, что на всех стадиях развития под пологом сероолыпаника обильно развивается порослевой подрост ольхи (в 60-летней синузии его плотность достигает 28000 шт./га). Накопление листового опада в подстилке и сильное затенение препятствуют прорастанию семян ели и развитию ее подроста, в тоже время устойчиво и обильно развивается подрост ольхи. Соответственно можно сделать вывод, что в сероолыпаниках наблюдается блокировка автогенной сукцессии, направленной
на формирование коренного типа сообщества -ельника.
В ходе развития березняка, осинника и серо-олыпаника наблюдаются существенные изменения в напочвенном покрове. За счет снижения освещенности, а в случае ольшаника – и увеличения обилия листового опада и накопления подстилки -происходит подавление развития травянистой растительности: достоверно снижается общее проективное покрытие, покрытие трав. Анализ влияния березы, осины и ольхи на совокупность всех видов напочвенного покрова (отношение суммы фактори-альных дисперсий всех видов под влиянием изучаемой породы к сумме общих дисперсий) показал, что наиболее слабое воздействие оказывает береза, а наиболее сильное – ольха (табл. 2).
Что касается изменения видового разнообразия и обилия отдельных видов, то прослеживается некоторая разнохарактерность влияния березы, осины и ольхи на эти показатели. Так, ольховая синузия негативно влияет на видовое разнообразие (табл. 2). Среднее число видов на площадке в 0,1 м2 падает с 13 на лугу до 2-х под пологом ольхи; для 70 % видов обнаружено достоверное отрицательное влияние, и только 10 % видов демонстрируют по-
ложительную связь. Особенно негативное воздействие оказывает развивающийся ольшаник на ге-лиофитные виды лугов: Phleum pratense (т)2=(-)0,30), Роа pratensis (г|2=(-)0,53), Deschampsia cespitosa (т)2=(-)0,44у), Centaurea jacea (rj2=(-)0,59), Ranunculus acris (rj2=(-)0,37), Lathyrus pratensis (r|2=(-)0,29). Анализ линий регрессии эколого-морфологических групп видов показывает, что обилие видов лугового разнотравья и граминоидов (злаков, осок, ситников) неуклонно падает, достигая нулевой отметки в 60-летнем ольшанике, а лесные виды, появившись под кроной ольхи, по ходу продвижения вглубь ольховой синузии, увеличивают свое участие: Oxalis acetosella (r|2=0,45), Moehringia trinervia (т)2=0,42). Семигелиофитные виды лугового крупно-травья (Anthriscus sylvestris, Filipéndula ulmaria, Angélica sylvestris) наиболее обильны на краю луга в зоне притенения ольхой. Здесь за счет снижения обилия злаков, уменьшается плотность луговой дернины и для этих видов складываются оптимальные условия. Аналогичным образом ведут себя зеленые мхи – представители рр. Plagiomnium и Aulacomnium. В зоне развитого ольхового древостоя их появлению препятствует обильный листовой опад, а на лугу – мощная злаковая дернина.
Таблица 2. Влияние березы, осины и ольхи на параметры растительности и биотопа
Береза Осина Ольха
Параметры г]2 г г]2 г г]2 г
Общее проективное покрытие 0,64 -0,35 0,34 -0,45 0,82 -0,82
Проективное покрытие трав 0,66 -0,28 0,35 -0,47 0,78 -0,78
Проективное покрытие мхов 0,02 0,05 0,12 0,25 0,40 -0,44
Число видов / 0,1 м2 0,54 -0,31 0,01 -0,09 0,86 -0,88
Совокупность видов 0,13 0,38 0,70
Проективное покрытие опада 0,82 0,51 0,78 0,73 0,94 0,93
Проективное покрытие ветоши 0,78 -0,44 0,89 -0,81 0,85 -0,91
Толщина подстилки 0,71 -0,50 0,45 -0,42 0,65 0,71
Примечание: ц2 – квадрат корреляционного отношения, влияющий фактор – зона (возраст зарастания); г – коэффициент корреляции Пирсона; значения, достоверные на уровне значимости 0,95, выделены жирным шрифтом.
В березняке по сравнению с лугом видовое разнообразие снижается в два раза (среднее число видов на площадке на лугу – 13, в березняке – 6). Отрицательное влияние березы выявлено для 31 % видов – это гелиофитные луговые виды: Festuca pratensis (rj2=(-)0,24), Phleum pratense (rj2=(-)0,17), Lathyrus pratensis (rj2=(-)0,18), Ranunculus acris (r)2=(-)0,20), Galium album (r)2=(-)0,ll) и др. 20 % видов демонстрируют положительную связь. Было показано ранее, что эти виды на суходольных лугах имеют между собой положительные связи и проявляют тип взаимоотношения «благоприят-ствование» [3]. Травянистый ярус березняка складывается преимущественно из группы лесных видов при малой доле участия луговых, для которых изменившиеся условия оказались благоприятными, например, для Deschampsia cespitosa, Angelica sylvestris, Melampy-rum nemorosum. Анализ линий регрессии подтверждает все выше сказанное: обилие лугового мелко-
травья и граминоидов неуклонно падает. Виды еловой свиты в березняке пока еще не показывают высокой встречаемости, но на отдельных площадках уже достигают значительных покрытий: Руг ola го-tundifolia (N=0,12; Smax=45 %), Majanthenum Ы-folium (N=0,10; Smax=35 %), Trientalis europaea (N=0,04; Smax=10 %), Paris quadrifolia (N=0,11; Smax=20 %).
В осиннике по сравнению с лугом, в отличии с ольшаника и березняка, достоверного снижения видового разнообразия не обнаружено (табл. 2) (среднее число видов на площадке на лугу – 11, в осиннике – 10). Отрицательное влияние осины выявлено лишь для 25 % видов – это все те же гелиофитные луговые виды. 38 % видов демонстрируют положительную связь, как и в случае с ольхой это лесные виды, но их разнообразие и обилие в осиннике значительно выше. В осиннике не формируется мощная подстилка как в ольшанике, что дает воз-
можность на фоне умеренного затенения успешно развиваться как большему числу лесных кустарничков и трав (Oxalis acetosella Rubus saxatilis (r|2=0,42), Trientalis europaea (г^О.Зб). Vaccinium vitis-idaea (г^О.ЗЗ)). так и зеленым мхам (табл. 2). Снижение обилия демонстрируют граминоиды, луговое крупнотравье и мелкотравье, но в отличие от ольшаника, эти группы видов не исчезают, а сохраняются на всех стадиях развития осинника, изменяя при этом свой видовой состав. Наиболее кардинально меняется видовой состав граминоидов: на смену луговым видам, таким как Alopecuruspratensis (r)2=(-)0,74), Phleum pratense (ri?=(-)0,26), Роа pratensis (г|2=(-)0,18), приходят лесные и опушечные виды: Calamagrostis canescens (г|2=0,40), С. purpurea 0f=0,44).
Изменения флористического и ценотического состава в ходе зарастания луговых фитоценозов мелколиственными породами попытались проиллюстрировать анализом коэффициентов флористического сходства Съеренсена (Ks) и ценотического сходства Глизона (KG). В ходе сравнения растительности луга и 60-летнего березняка были зафиксированы самые высокие значения коэффициентов сходства: Ks=65 %, KG=47 %. В случае с осинником эти показатели ниже: Ks=45 %, KG=21 %. И самые низкие значения коэффициентов сходства наблюдаются при сравнении луга и 60-летнего серооль-шаника: Ks=15 %, Kg=8 %. Низкие значения этих коэффициентов свидетельствуют о смене растительных сообществ, что особенно явственно.
Таким образом, под пологом развивающихся древостоев березы, осины и ольхи серой существенно преображается биотоп: снижается освещенность, изменяется температурный режим и влажность, преобразуется характер и обилие подстилки, снижается доля участия луговых видов вплоть до их полного исчезновения и возрастает покрытие лесных видов.
В случае зарастания лугов ольхой серой наблюдается резкое снижение видового богатства и обилия травяного покрова, мощная подстилка из листового опада блокирует развитие елового подроста, в результате наблюдается торможение прохождения дальнейших стадий автогенной сукцессии, что не происходит при развитии березовых и осиновых синузий.
Подводя итоги, можно сделать следующие выводы. Зарастание лугов мелколиственными породами сопровождается изменением биотопа, преобразованием состава и структуры расти-тельных сообществ и приводит к смене лугового типа растительности лесным. Проведенный анализ интенсивности влияния трех мелколиственных пород на преобразование растительного покрова позволил выявить следующий ряд увеличения эдификаторной силы: береза -осина – ольха серая. Дальнейшее беспрепятственное зарастание луговых фито-ценозов мелколиственными древесными породами – березой, осиной и ольхой серой – на территории Нижне-Свирского государственного природного заповедника может привести к исчезновению луговой растительности и снижению биологического разнообразия на видовом, популяционном и ценотическом уровнях.
Работа выполнена при поддержке Российского фонда фундаментальных исследований.
СПИСОК ЛИТЕРАТУРЫ
1. Василевич В.И., Бибикова Т.В. Щучковые и лисохво-стные луга северо-запада Европейской России // Бот. журн. 2007. Т. 92. №1. С. 29-41.
2. Ипатов В. С., Кирикова Л. А. Применение дисперсионного анализа при исследовании связи растительности со средой//Бот. журн. 1977. Т. 62. №10. С. 1441-1445.
3. Лебедева В. X., Тиходеева М. Ю., Ипатов В. С. К вопросу о структуре лугового растительного сообщества // Бот. журн. 2021. Т. 96. №1. С. 3-21.
THE RESTORATION OF WOOD VEGETATION ON PLACE OF MEADOWS © 2021 M.Y. Tikhodeeva, V.Ch. Lebedeva
St-Petersburg State University
The comparative analysis of changes in biotope, composition and structure of the upland meadows in growing their small-leaved species: the white birch (Betula pubescens), aspen (Populus tremula), grey alder (Alnus incana) is carried out; built increase the number of edificator powers of the trees is defined.
Keywords: structure of communities, habitat, edificator, meadows, woods
Tikhodeeva Marina Yurievna, Ph. D., leading researcher, e-mail: marinaur@list.ru; Lebedeva Vera Christoforovna, Ph. D., leading researcher, e-mail: vera_christ@mail.ru
Эколого-фитоценотическая характеристика суходольных лугов калининградской области
СПИСОК ЛИТЕРАТУРЫ
1. Абрамова Л.М., Миркин Б.М. Эволюция растительности на стыке тысячелетий // Теоретические проблемы экологии и эволюции. Третьи Любищевские чтения. Тольятти, 2000. С. 15-23.
2. Агроклиматические ресурсы Литовской ССР и Калининградской области РСФСР / отв. ред. П.И. Егорова, Ф.Ш. Чечик. Л., 1972. 144 с.
3. Александрова В. Д. Классификация растительности. Л., 1969. 275 с.
4. Алексеев Ю.Е. Определитель осок средней полосы Европейской части СССР по вегетативным признакам. М., 1971. 80 с.
5. Алексеев Ю.Е. Осоки: морфология, биология, онтогенез, эволюция. М., 1996. 251 с.
6. Алексеенко Л.Н. Продуктивность луговых растений в зависимости от условий среды. Л., 1967. 168 с.
7. Алексеенко Л.Н. Водный режим луговых растений в связи с экологическими условиями: автореф. дисс… докт. биол. наук. Л., 1976. 46 с.
8. Алехин В.В. Растительность СССР в основных зонах. М.-Л., 1936. С.306-694.
9. Алещенко Г.М., Букварева E.H., Щербаков A.B. Использование экспертных оценок для анализа территории по критериям биоразнообразия//Успехи соврем, биол. 1995. Т. 115. С. 645-654.
10. Андреев Н.Г. Луговодство. М., 1981. 383 с.
11. Андронова H.H., Кученева Г.Г. Флора Калининградской области // Изученность природных ресурсов Калининградской области. Зап. Калининград, отд. Географ, общества. Вып. 1. Л., 1972. С. 141-160.
12. Анциферова O.A. Тенденции современной эволюции залежных агролапдшафтов Калининградского полуострова // Проблемы сельского хозяйства: сб. науч. тр. Калининград, 2000. С. 81-88.
13. Анциферова O.A. Направленность и интенсивность изменений свойств почв на молодых залежах Замландского полуострова // Актуальные
14
15
16
17
18
19
20,
21,
22,
23,
24,
25,
26,
27.
проблемы сельского хозяйства: сб. науч. тр. Калининград, 2001. С. 164-173.
Анциферова O.A. Почвы Замландского полуострова. 4.1: Факторы почвообразования. Почвы подзолистого и буроземного рядов. Калининград, 2008. 397 с.
Бавтуто Г.А., Еремин В.М. Ботаника: Морфология и анатомия растений. Минск, 1997. С. 330-334.
Базилевич Н.И. Особенности функционирования травяных экосистем в сравнении с лесными и пустынными // Математическое моделирование в экологии. М., 1978. С. 65-100.
Базилевич Н.И., Гребенщиков О.С., Тишков A.A. Географические закономерности структуры и функционирования экосистем. М., 1986. 296 с.
Баринова Г.М. Калининградская область. Климат. Калининград, 2002. 196 с.
Бедарев С.А., Бедарева О.М. Научные основы организации исследований по экологическому мониторингу растительного покрова // Проблемы сельского хозяйства: сб. науч. тр. КГТУ. Калининград, 1999. С. 3-9.
Белолюбцев А.И., Суховеева О.Э., Асауляк И.Ф. Агроклиматическая оценка продуктивности озимой пшеницы па склоновых землях // Известия ТСХА, 2021. Вып. 2. С. 46-56.
Биологическая флора Московской области. М., 1976. Вып. 3. 182 с. Биологическая флора Московской области. М., 1980. Вып. 5. 192 с. Биологическая флора Московской области. М., 1990. Вып. 8. 272 с. Биологическая флора Московской области. М., 1995. Вып. 10. 208 с. Биологическая флора Московской области. М., 1996. Вып. 12. 182 с. Благовещенский Г.В. Европейская федерация луговодов (ЕФЛ) // Кормопроизводство. 2021. № 1. С. 8-10. Будыко М.И. Изменения климата. Л., 1969. 38 с.
28. Будыко М.И. Климат и жизнь. JI.,1971. 472 с.
29. Будыко М.И.Эволюция биосферы. Л., 1984. 488 с.
30. Василевич В.И. Доминанты в растительном покрове // Ботанический журнал. 1991. Т. 76, № 12. С. 1674-1681.
31. Виноградова О.Л. Аллювиальные почвы – возможный объект-индикатор состояния ландшафтов бассейна р. Преголи // Проблемы сельского хозяйства: сб. науч. тр. Калининград, 2000. С.76-80.
32. Виноградова О.Л. Аллювиальные почвы Калининградской области и их хозяйственное использование // Проблемы сельского хозяйства: сб. науч. тр. Калининград, 1999. С.111-114.
33. Воробьева Л.А. Химический анализ почв. М., 1998. 271 с.
34. Географический атлас Калининградской области / гл. ред. В.В. Орленок. Калининград, 2002. 276 с.
35. География янтарного края России / под ред. В.В. Орленка. Калининград, 2004. С. 97.
36. Герасимова М.И., Гаврилова И.П. Автономные суглинистые почвы центральной части Калининградской области: проблемы генезиса // Почвоведение. 2005. №1. С. 5-15.
37. Гогмачадзе Г.Д. Агроэкологический мониторинг почв и земельных ресурсов Российской Федерации. М., 2021. 592 с.
38. Горчаковский П.Л. Антропогенные изменения растительности: мониторинг, оценка, прогнозирование//Экология. 1984. № 4. С. 3-16.
39. Горчаковский П.Л. Основы охраны и экологического мониторинга луговой растительности // Продуктивность сенокосов и пастбищ. Новосибирск, 1986. С. 25-28.
40. Горчаковский П.Л. Антропогенная трансформация и восстановление продуктивности луговых фитоценозов. Екатеринбург, 1999. 156 с.
41. Горчаковский П.Л., Абрамчук A.B. Формирование и деградация суходольных лугов под влиянием выпаса и сенокошения // Экология. 1993. №4. С. 3-13.
42. Горчаковский ПЛ., Телегова О.В. Сравнительная оценка уровня синантропизации растительного покрова особо охраняемых природных территорий // Экология. 2005. № 6. С. 403-408.
43. Горышина Т.Г. Экология растений. М., 1979. 367 с.
44. ГОСТ 26483-85 Почвы. Приготовление солевой вытяжки и определение ее рН по методу ЦИНАО. URL: http://docs.cntd. ru/document/1200023490 (дата обращения: 15.01.10).
45. ГОСТ 27821-88 Почвы. Определение суммы поглощенных оснований по методу Каппена. URL: http://docs.cntd.ru/document/1200023552 (дата обращения: 20.01.10).
46. ГОСТ 26212-91 Почвы. Определение гидролитической кислотности по методу Каппена в модификации ЦИНАО. URL: http://docs.cntd. ru/document/ 1200023469 (дата обращения: 22.05.10).
47. ГОСТ Р 54650-2021 Почвы. Определение подвижных соединений фосфора калия по методу Кирсанова в модификации ЦИНАО. URL: http://docs.cntd. ru/document/ 1200094361 (дата обращения: 27.07.11).
48. Губанов И.А. Луговые травянистые растения. Биология и охрана. М., 1990. С. 1-15.
49. Губанов И.А, Киселева К.В., Новиков B.C., Тихомиров В.Н. Gramineae (Роасеае) – Злаковые // Иллюстрированный определитель растений Средней России. М., 2002. Т. 1. С. 181-313.
50. Гулинова Н.В. Погода и урожай сеяных и луговых трав. Л., 1982. 174 с.
51. Гулинова Н.В. Агрометеорологические условия, баланс кормов и продуктивность животноводства. Л., 1988. 247 с.
52. Дедков В.П., Семенова З.Е., Губарева И.Ю. и др. Охраняемые растения и растительные сообщества Калининградской области / под ред. В.П. Дедкова. Калининград, 1990. 88 с.
53. Дедков В.П., Губарева И.Ю., Петрова Н.Г. и др. Современное состояние изученности флоры Калининградской области // XXVIII
науч. конф. Калннннгр. гос. ун-та: Тез. докл. Ч. 1. Калининград, 1997. С. 50-51.
54. Диагнозы и ключи возрастных состояний злаков. Методические особенности для студентов биол. специальностей. М., 1997. 141 с.
55. Дмитриев С.И., Игловиков В.Г., Кошошков Ы.С. и др. Растения сенокосов и пастбищ / под ред. С И. Дмитриева. М., 1982. 195 с.
56. Еремин Т.П., Беднова Е.Т. Состояние и меры улучшения лугов и пастбищ Калининградской области. Калининград, 1949. 56 с.
57. Елисеева И.И. Статистика. СПб., 2021. 368 с.
58. Ермолаев A.M., Ширшова JI.T. Продуктивность и функционирование многолетнего сеяного луга различного режима использования // Почвоведение. 1994. № 12. С. 97-105.
59. Ерохип В.В. Флора и растительность суходольных лугов правобережья реки Вятки в нижнем ее течении: Автореф. дис. … канд. биол. наук. Казань, 2003. 20 с.
60. Жмылев П.Ю., Алексеев Ю.Е., Карпухина В.А. Биоморфология растений: иллюстрированный словарь. М., 2002. 256 с.
61. Жукова JI.A. Методология и методика определения экологической валентности, стеноэврибионтности видов растений // Методы популяционной биологии: сб. материалов VII Всерос. популяционного семинара. Сыктывкар, 2004а. 4.1. С. 75-76.
62. Жукова JI.A. Оценка экологической валентности основных эколого-ценотических групп: подходы и методы // Восточно-европейские леса: история в голоцене и современность. 20046. Кн. 1. С.256-259.
63. Жукова JI.A. Экологическое разнообразие ценопопуляций модельных видов растений в Национальном парке «Марий Чодра» // Биоразнообразие растений в экосистемах Национального парка «Марий Чодра». Йошкар-Ола, 2005. Ч. 2. С.19-32.
64. Жукова JI.A., Турмухаметова Н.В., Акшепцев Е.В. Экологическая характеристика некоторых видов растений // Онтогенетический атлас
растений. Йошкар-Ола, 2007. Т.5. С. 318-331.
65. Зайкова В.А. Динамика луговых сообществ. JI., 1980. 216 с.
66. Завалишин A.A., Надеждин Б.В. Почвенный покров Калининградской области//Почвы Калининградской области. М., 1961. С. 5-130.
67. Заугольнова Л.Б., Истомина И.И, Тихонова Е.В. Экологический, ценотический и флористический анализ растений лесной зоны европейской России на основе экологических шкал, геоботанических описаний и статистического анализа // Бюл. МОИП. Сер. Биологическая. 2001. Т. 111. №2. С. 36-47.
68. Зозулин Г.М. Исторические свиты растительности Европейской части России //Ботанический журнал. 1973. Т. 58, № 8. С. 1081-1092.
69. Исаченко А.Г. Ландшафтоведение и физико-географическое районирование. М., 1991. 365 с.
70. Ипатов B.C. Оценка данных при проективном учете // Ботанический журнал. 1964. Т. 49, № 3. С. 382-386.
71. Ипатов B.C. Описание фитоценоза. СПб, 1998. 151 с.
72. Итоги 45-летнего изучения Залидовских лугов Калужской области. Памяти профессора A.A. Уранова. // Е.И. Курченко, И.М. Ермакова, Н.С. Сугоркина, В.Г. Петросян, Ф.А. Маслов. Материалы Всерос. науч. копф. «Отечественная геоботаника: основные вехи и перспективы. СПб., 2021. Т.2. С. 125-128.
73. Калинина С.И., Лайдинен Г.Ф. Биологические основы возделывания многолетних злаковых трав на европейском севере России. Петрозаводск, 1995. 212 с.
74. Калининградская область: очерки природы / сост. Д.Я. Беренбейм. Калининград, 1999. 229 с.
75. Клапп Э. Сенокосы и пастбища. М., 1961. 613 с.
76. Классификация и диагностика почв СССР / сост. В.В. Егоров, В.М. Фридланд, E.H. Иванова и др. М., 1977. 224 с.
77. Климов Г.П. Теория вероятности и математическая статистика. М.,
2021. 368 с.
78. Козловская H.B. Флора Белоруссии: закономерности ее формирования, научные основы использования и охраны. Минск, 1978. 128 с.
79. Кормовые растения сенокосов и пастбищ СССР. M.-JI, 1950. Т.1. 649 с.
80. Кормовые растения сенокосов и пастбищ СССР. M.-JI, 1951. Т. 2. 988 с.
81. Котелина Н.С. Ядовитые растения сенокосов и пастбищ. Сыктывкар, 1973. 172 с.
82. Кочуров Б.И., Розанов Л.Л. Районирование территории России по современной экологической ситуации / Региональные аспекты развития России в условиях глобальных изменений природной среды и климата. М., 2001. С. 56-64.
83. Кузнецова Л.В., Нешатаев В JO. Проблема классификации мезофитнотравяных урбанофитоцепозов // Мат-лы Всерос. науч. конф. «Отечественная геоботаника: основные вехи и перспективы. СПб., 2021. Т. 1.С. 127-129.
84. Куркин К.А. О филоценогенезе и селектоценогенезе в связи с биохимической эволюцией ландшафтов // Теоретические проблемы фитоценологии и биогеоценологии. М., 1970. С. 61-73.
85. Куркин К.А. Системные исследования динамики лугов. М., 1976. 284 с.
86. Куркин К.А. Опыт экологической классификации растительности пойменных лугов. Обоснование ландшафтно-зкологической классификации (по частям поймы) // Ботанический журнал. 1987. Т. 72, № 12. С. 1605-1616.
87. Куркин К.А. Экологические факторы дифференциации луговой растительности //Ботанический журнал, 1992. Т. 77, № 6. С. 30-41.
88. Куркин К.А. Луговой тип растительности и его отграничение от других типов // Ботанический журнал, 1996. Т. 81, № 8. С. 12-26.
89. Кутузова A.A., Трофимова Л.С., Орленкова Е.К. Продуктивность
долголетних сенокосов при разных режимах // Кормопроизводство. 2001. №9. С. 10-14.
90. Кучерявый П.П. География Калининградской области / П.П. Кучерявый, Г.М. Федоров. Калининград, 1989. 142 с.
91. Ларин И.В., Матвеева Е.П., Сырокамская И.В. Динамика развития луговой растительности Калининградской области // Тр. БИН АН СССР. 1955. Сер. 3. Вып. 10. С. 31-101.
92. Малышев Л.И. Оценка оригинальности флоры по таксономической структуре // Ботанические исследования Сибири и Казахстана. 2000. Вып. 6. С. 3-10.
93. Мартыненко В. А. Флористический состав кормовых угодий европейского Северо-Востока. Л., 1989. 135 с.
94. Маракулина C.IO. Суходольные луга таежной зоны Кировской области. Автореф. дис. … канд. биол. наук. Сыктывкар, 2009. 21 с.
95. Матвеева Е.П. Сенокосы и пастбища и меры их улучшения (из материалов Калининградской экспедиции). М.-Л., 1953. 110 с.
96. Матвеева Е.П. Луга советской Прибалтики, Л., 1967. 335 с.
97. Методика опытных работ на сенокосах и пастбищах. / Под ред. Н.С. Кошошкова, Т.А. Работнова, И.А. Цаценкина. М., 1961. 287 с.
98. Миняев H.A. Флористическое районирование северо-запада европейской части СССР // Тр. Лепингр. о-ва естествоиспыт. 1959. Т. 70, вып. 1.С. 35-37.
99. Миняев H.A. Об арктических и альпийских связях во флоре северо-запада европейской части СССР // Вопросы ботаники. 1960. Вып. 3. С. 63-65.
100. Миняев Н. А. Арктические и арктоальпийские элементы во флоре северо-запада европейской части СССР // Ареалы растений флоры СССР. Л., 1965а. С. 9-49.
101. Миняев H.A. Сибирские таежные элементы во флоре северо-запада европейской части СССР //Ареалы растений флоры СССР. Л., 19656.
С. 50-92.
102. Миняев II. А., Ниценко А. А. Состояние и организация охраны растительных объектов Северо-Запада европейской части СССР // Вопросы охраны ботанических объектов. JL, 1971. С. 206-212.
103. Миркин Б.М., Розенберг Г.С. Фитоценология. Принципы и методы. М., 1978. 212 с.
104. Миркин Б.М., Розенберг Г.С. Количественные методы классификации, ординации и геоботапической ординации // Итоги науки и техники. Ботаника. 1979. Т.2, С. 71-137.
105. Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М., 2001. 264с.
106. Миркин Б.М., Мартыпенко В.Б., Л.Г. Наумова Значение классификации растительности для современной экологии // Журнал общей биологии. 2004. Т. 64, № 2. С. 167-177.
107. Морозова О.В. Таксономическое богатство флоры Восточной Европы: факторы пространственной дифференциации. М., 2008. 328 с.
108. Наумова Л. Г. Основы фитоценологии. Уфа, 1995. 238 с.
109. Нешатаев Ю.Н. Методы анализа геоботанических материалов. Л., 1987. 192 с.
110. Никитин В.В. Сорные растения флоры СССР. Л., 1983. 455 с.
111. Ниценко A.A. Об изучении экологической структуры растительного покрова //Ботанический журнал, 1969. Т. 54, № 7. С. 1002 – 1014.
112. Ниценко A.A. Растительные ассоциации и растительные сообщества как первичные объекты геоботанического исследования. Л., 1971. 184 с.
113. Носова Л.М. Флоро-географический анализ северной степи европейской части СССР. М., 1973. 187 с.
114. Определитель высших растений Северо-Запада европейской части РСФСР (Ленинградская, Псковская и Новгородская области)/Сост. H.A. Миняев, Н.И.Орлова, В.М. Шмидт и др. Л., 1981. 376 с.
115. Орленок B.B. Региональная география России. Калининградская область / В.В. Орленок, Г.М. Федоров. Калининград, 2005. 259 с.
116. Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / О.В. Смирнова, Л.Б. Заугольнова, Л.Г. Ханина и др. М., 2000. 196 с.
117. Павлова Г.Г. Суходольные луга правобережья Енисея: автореф. дис. … канд. биол. наук. Новосибирск, 1973. 28 с.
118. Павлова Г.Г. Суходольные луга юга Средней Сибири. Новосибирск, 1980.216 с.
119. Панасин В.И. Агрохимические свойства почв Калининградской области и применение удобрений. Калининград, 1974. 272 с.
120. Панасин В.И. Микроэлементы и урожай. Калининград, 1994. 282 с.
121. Панасин В.И., Слободжанипова В.Д., Новикова С.М. Агрохимическая характеристика почв Калининградской области. Калининград, 1997. 79 с.
122. Панасин В.И., Брысозовский И.И., Слободжанипова В.Д. и др. Медь и урожай. Калининград, 1999. 240 с.
123. Панасин В.И. Микроэлементы и урожай. Калининград, 2000. 276 с.
124. Панасин В.И. Гумус и плодородие почв Калининградской области. Калининград, 2004. 220 с.
125. Паракшина Э.М., Терентьева М.Ю. Итегративная эрозия почв в агроландшафтов Русской равнины // Актуальные проблемы сельского хозяйства: сб. науч. тр. Калининград, 2001. Ч. 1. С. 77-88.
126. Плаксина Т.И. Флора Волго-Уральского региона: автореф. дис. … д-ра биол. наук. М., 1994. 36 с.
127. Плаксина Т.И. Конспект флоры Волго-Уральского региона. Самара, 2001.388 с.
128. Победимова Е.Г. Состав, распространение по районам и хозяйственное значение флоры Калининградской области / Труды Ботанического инта АН СССР, серия III. Л, 1956. 325 с.
129. Понятовская В.М. Учет обилия и особенности размещения видов в естественных растительных сообществах // Полевая геоботаника. М.-Л., 1964. Т. 3. С. 209-299.
130. Природные сенокосы и пастбища / под ред. И.В. Ларина. М.-Л., 1963. 548 с.
131. Пузаченко Ю.Г. Математические методы в экологических и географических исследованиях. М., 2004. 416 с.
132. Работнов Т.А. Что считать лугом? // Ботанический журнал. 1959. Т. 44, № 1.С. 35-43.
133. Работнов Т.А. Изменчивость луговых ценозов и ее значение для практики геоботанических исследований // Геоботаническое изучение лугов: сб. бот. работ Белорус, отд. ВБО. Вып. 4. Минск, 1962. С. 5-17.
134. Работнов Т.А. Суходольный луг как биогеоценоз. М., 1978. 84 с.
135. Работнов Т.А. Луговедение. М., 1984. 319 с.
136. Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л., 1971. С. 245-247.
137. Расширенная система эколого-ценотических групп видов сосудистых растений для бореальной, гемибореальной и умеренной лесных зон Европейской России: [сайт ИМПБ РАН]. URL: http://www. impb.ru/spm/geobot./ecg (дата обращения: 10.03.2021).
138. Сагалаев В.А. Флора степей и пустынь Юго-Востока Европейской России, ее генезис и современное состояние: дис … д-ра биол. наук. М., 2000. 1005 с.
139. Сапожникова С.А., Шашко Д.И. Агроклиматические условия размещения и специализация сельскохозяйственного производства. Л., 1959.25 с.
140. Селянинов Г.Т. Методика сельскохозяйственной характеристики климата / Мировой агроклиматический справочник. Л.-М., 1937. С. 527.
141. Семенова-Тян-Шанская A.M. Накопление и роль подстилки в
травяных сообществах. Л., 1977. 191 с.
142. Серебряков И.Г. Морфология вегетативных органов высших растений. М., 1952. 392 с.
143. Серебряков И.Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных растений. М., 1962. 378 с.
144. Серебряков И.Г. Жизненные формы высших растений и их изучение // Полевая геоботаника. М.-Л., 1964. Т. 3. С. 146-205.
145. Серебрякова Т.И. Учение о жизненных формах растений на современном этапе // Итоги науки и жизни. М., 1972. Т.1. С. 84-169.
146. Смирнов В.Э., Ханина Л.Г., Бобровский М.В. Основание системы эколого-ценотических групп видов групп ассоциаций хвойно-широколиственных лесовцентра Европейской России // Растительность России. 2006. №7. С. 38-48.
147. Сохранение и восстановление биоразнообразия / под ред. В.Е. Флинт. М., 2002. 286 с.
148. Справочник по кормопроизводству / сост. В.Г. Игловиков, Н.С. Кошонжов, А.П. Мовсисянц и др. М., 1973. 488 с.
149. Справочник по кормопроизводству / сост. А.И. Тютюнников. М., 1982. 352 с.
150. Справочник по сенокосам и пастбищам /сост. Д.А. Алтунин. М., 1986. 355 с.
151. Степанович И.М. Мониторинг луговой растительности // Национальная система мониторингаокружающей среды Республики Беларусь: результаты наблюдений. Минск, 2005а. С.133-146.
152. Степанович И.М., Ивкович E.H., Степанович Е.Ф. Автушко С.А. Травяные сообщества Березинского биосферного заповедника: структура, продуктивность, состояние. Минск, 20056. 200 с.
153. Степанович И.М., Степанович Е.Ф. Система мониторинга луговой и лугово-болотной растительности Беларуси // Мат-лы Всерос. науч. конф. «Отечественная геоботаника: основные вехи и перспективы.
СПб., 2021. Т.2. С. 127-129.
154. Стратегия сохранения редких и находящихся под угрозой исчезновения видов животных, растений и грибов: приказ министерства природных ресурсов РФ от 6 апреля 2004 г. №323. Доступ из справ.-правовой системы «КосультантПлюс».
155. Схема охраны природы Калининградской области / под ред. Ю.А. Цыбина. Калининград, 2004. 136 с.
156. Тахтаджян A.JI. Флористические области Земли. Д., 1978. 247 с.
157. Тахтаджян A.JI. Флористическое деление суши / А Л. Тахтаджян // Жизнь растений. М. 1980-1982. Т. 5(1), Т. 5(2), Т 6.
158. Титлянова A.A. Продуктивность травяных экосистем // Биологическая продуктивность травяных экосистем. Новосибирск, 1988. С. 109-127.
159. Толмачев А.И. Введение в географию растений. JL, 1974. 244 с.
160. Трофимова Л.С., Трофимов И.А., Яковлева И.П. Агроландшафтно-экологическое районирование кормовых угодий Северо-Западного природно-экономического района РФ // Кормопроизводство. 2021. № 8.С. 10-13.
161. Утехин В.Д. Первичная биологическая продуктивность лесостепных экосистем. М., 1977. 144 с.
162. Федоров Е.Е., Галахов H.H., Гальцов А.П. Климат Калининградской области // Почвы Калининградской области. М., 1981. С. 159-174.
163. Филатова Т.Д. Растительность залежи «Дальнее поле» при разных режимах заповедания (Казацкий участок Центрально-Черноземного заповедника) // Флора и растительность Центрального Черноземья. Курск, 2003. С. 64-66.
164. Флора Северо-Востока европейской части СССР. Л., 1974. T. I. 257 с.
165. Флора Северо-Востока европейской части СССР. Л., 1976а. Т. П. 316 с.
166. Флора Северо-Востока европейской части СССР. Л., 19766. Т. III. 293 с.
167. Флора Северо-Востока европейской части СССР. Л., 1977. T. IV. 312 с.
168. Хохряков А.П. Таксономические спектры и их роль в сравнительной флористике // Ботанический журнал. 2000. Т. 85, № 5. С. 1-11.
169. Цаценкин И.А. Типологический состав естественных сенокосов и пастбищ // Вестник сельскохозяйственной науки. М., 1974. С.95-107.
170. Цвелев H.H. Злаки СССР. Л., 1976. 322 с.
171. Цвелев H.H. Семейство Злаки – Роасеае // Систематика растений: Однодольные растения. Семейства ситниковые, осоковые, злаки: Учебное пособие. Ч. 2. СПб., 1994. С. 46-90.
172. Цвелев H.H. Определитель растений Северо-Западной России (Ленинградская. Псковская и Новгородская области). СПб., 2000. 781 с.
173. Целевая Программа Калининградской области «Развитие мясного скотоводства в Калининградской области на 2021-2021 годы» от 5 декабря 2021 г. № 914. Доступ из справ.-правовой системы «КосультаптПлюс».
174. Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М., 1983. 197 с.
175. Черепанов С.К. Сосудистые растения России и сопредельных государств. СПб., 1995. 990 с.
176. Шенников А.П. Принципы ботанической классификации лугов // Сов. ботаника. 1935. №5. С. 35-49.
177. Шенников А.П. Луговая растительность СССР // Растительность СССР. М.-Л., 1938. Т. 1. С. 429-647.
178. Шенников А.П. Луговедение. Л., 1941. 512с.
179. Шенников А.П. Экология растений. М., 1950. 375 с.
180. Шенников А.П. Изучение лугов СССР // Вопр. ботаники. М., 1954. Вып. 1.С. 363-386.
181. Шереметьева И.С. Флора Тульской области: дис …канд. биол. наук. М, 1999. 455 с.
182. Шилов М.П., Уразов И.А. Охрана лугов. Иваново, 1984. 95 с.
183. Шилов М.П. Состояние, использование, оптимизация и охрана естественных сенокосов и пастбищ Нечерноземной зоны РСФСР // Продуктивность сенокосов и пастбищ. Новосибирск, 1986. С. 85-96.
184. Экологическая оценка кормовых угодий по растительному покрову / Л.Г.Раменский, H.A. Цаценкин, О.Н. Чижиков, H.A. Антипин. М., 1956. 472 с.
185. Экологические шкалы и методы анализа экологического разнообразия растений / под общ. ред. Л.А Жуковой. Йошкар-Ола, 2021. 368 с.
186. Юрцев Б.А. Мониторинг биоразнообразия на уровне локальных флор // Ботанический журнал. 1997. Т.82, № 6. С. 60-69.
187. Abromeit J. Flora von Ost-und Westpreussen. – Königsberg, 1898-1940.
188. Abromeit J., Neuhoff W., Steffen H., Jentzsch A., Vogel G. Flora von Ost-und Westpreussen. Bd. Berlin, 1903.
189. Awater S. Gräser und Krauter, 2006, Bonn, 12 s.
V
190. Daugeliene N. Ganykliniai zolynai, jq derlingumas ir pasarine verte / Zemes Okio mokslai. Vilnius, 1995a. 124 psl.
V
191. Daugeliene N. Ganykliniai zolynai ir ji} atseliavimas / LZI mokslo darbai. Dotnuva – Akademija, 1995b. t. 43. 90 psl.
192. Dierschke PI. Pflanzensoziologie. Grundlagen und Methoden. Ulmer Verlag Stuttgart 1994. 682 s.
193. Dierschke PI., G.Briemle Wiesen, Weiden und verwandte Staudenfluren. Ulmer Verlag Stuttgart. 2002. 451 s.
194. Dierßen K. Hemerobiestufen des Feuchtgrundlandes in Sleswig-Holstein // Erf. und Bewert, antropogener Vegetationsveranderungen. Teil II. // Wiss. Beitr. Martin-Luter Univ.,1987. Bd. 24. S. 4-25.
195. Dierßen K. Vegetation Nordeuropas / Unter Mitarb. von K. Dierßen. Verlag Stuttgart (Hohenheim). 1996. 838 S.
196. Fagerberg В., Nyman P. The effect of weather fluctuations on simulated ley growth in Sweden / Swed.J. agr.Res., 1991. Vol. 21. № 3. P. 95-105.
197. Geißelbrecht-Taferner L., Mucina L. Vegetation der Brachen der Stadt
Linz. Linz: Staphia, 1995. Bd. 38. 154 S.
198. Haken D., Kvitek T. Vliv klimatickych podminek na kolisani vynosu luk a pastvin / CS Acad. Zemed. Praha. 1986. T. 107. S. 88-94.
199. Hutter C.-P., Briemle G., Fink C. Wiesen, Weiden und anderes Grünland. Weitbrecht Verlag 1993. 385 s.
200. Jager G. Möglichkeiten der Prognose synantroper Pflanzenausbreitungen // Flora. 1988. № 180. S. 101-131.
201. Klapp E. Zur Problematik des Ackers-und Pflanzenbaues (Vortrag aus dem Jahre 1951). Verlag Adelheid Böhm, Göttingen, 1994. 476 s.
202. Nachhaltige Grünlandwirtschaftung in Baden-Württenberg von G.Briemle, M. Elsäßer, T.Jilg, W.Müller, H.Nußbaum / Nachhaltige Land-und Forstwirtschaft. 1996. S.215-256.
203. Passarge II., Iloffman G. Pflanzengesellschaften des nordostdeutschen Flachlandes. Gustav Fischer Verlag Jena, 1968. S.3-298.
204. Pott R. Die Pflanzengesellschaften Deutschlands. Verlag Eugen Ulmer Stuttgart. 1992. 370 s.
205. Pott R., Hüppe J. Spezielle Geobotanik. Pflanze-Klima-Boden. SpringerVerlag, Berlin, Heidelberg, New York. 2007. 330 s.
206. Pott R., Küster I-I. Der Klimawandel und die Vegetation. Garten und Landschaft 2007. 117 (8). S.20-21
207. Runge F. Die Pflanzengesellschaften Deutschlands. Münster Verlag Aschendorff 1973.230 s.
208. Schlüter H. Vegetationsmerkmale zur Kennzeichnung und Bewertung von I-Iemerobie und ekologischer Stabiiitat // Erfass. Und Bewert. Antropogener Vegetationsveranderungen. Teil II. // Wiss. Beitr. Martin-Luter Univ., 1987. Bd. 25. S. 13-19.
209. Taschenbuch der Graser. Erkennung und Bestimmung.Standort und Vergesellschaftung. Von W.Opitz von Boberfeld überarbeitete E.Klapps Auflage. Verlag Eugen Ulmer, Stuttgart. 2006. 456 s.
210. Tüxen R. Grundfragen und Methoden in der Pflanensoziologie. Verlag
Dr.W.Junk N.V. 1972. 250 s.
211. Wiesen und Weiden. Eine Grünlandlehre. Überbearbeitet von mehreren Fachkollegen. Berlin, 1971. 480 s.
212. Wilde Weiden. Praxisleitfaden für Ganzjahresbeweidung in Naturschutz und Landschaftsentwicklung. M.Bunzel-Drüke, C.Böhm, P.Finck, G.Kämmer, R.Luick und andr. Arbeitsgemeinschaft Biologischer Umweltschutz im Freis Soest e.V. 2009. S.l 12-163.
213. Wilmanns O. Ökologische Pflanzensiziologie-eine Einfuhrung in die Vegetation Mitteleuropas Wiesbaden, 1998. 411 s.
214. Valionis E. Tik sveikas gyvulys naudingas. Kaunas, 1997. 112 psl.





