
Биологические и физические процессы и закономерности в живых системах. редукционизм и антиредукцианизм. принцип качественной несводимости.
Биологические и физические процессы и закономерности в живых системах. Редукционизм и антиредукцианизм. Принцип качественной несводимости.
Живые огранизмы – открытая, саморег-ся, самовоспр-ся и разв-ся гетерогенная система, важнейшими функциональными в-вами в которой явл-ся биополимеры: белки и нукл. к-ты сложного атомно-мол-ного строения.
Первые попытки объяснения биолог. пр-сов связаны с методами сравнения с физич. пр-ми. Напр: м-м нервного проведения – как распростр. волны окисления в медной проволоки в кислоте; мышечное сокращение объяснялось работой пьезо элементов; рост клетки. Первоначально физика проникла в химию – необходимость объяснить как разл. соединения взаимод-ют в организме – физическая химия и химическая физика.
Сущ 2 лагеря современ. физики:
1) Редукционисты: Любой био процесс происх. в жив организме можно свести к суме хим., физ. и механич. процессов. Объяснение сложного ч/з более простое, непонятное ч/з известное. Зная св-ва отдельных элементов, сост. систему и особенности их взаимод-вия, можно вывести все свойства этой системы. Обр. более сложного уровня – это результат усложнения более простых. Иногда: попытки заменить исследование реального объекта его упрощенной моделью. Достижение: предсказание существования планеты Нептун. Но как метод мышления не является универсальным. Неудача в биологии: не могут объяснить с этой т.зр. феномен жизни.
2) Антиредукционисты: Принцип качественной несводимости или био антиред., т.е. невозможность свести законы и принципы, управляющие живой материей к элементарной сумме физико-хим и мех. процессов процесс жизнедеятельности. Т.е. физико-матем. модели не м.б. адекватными, если в них не сод-ся элементы функциональной организации живых систем. Т.е. существует предел, после которого физические представления перестают быть самодостаточным средством познания, а дальше определяющим фактором становятся некоторые био истины, без которых уже не обойтись.
Основные направления развития современной биофизики. Уровни биофизических исследований.
Биофизика – это н., изуч физ и физико-хим. пр-сы, протекающие в биосис. на разных уровнях орг-ции и явл-ся основой физиолог-их актов.
Разделы биофизики: (и уровни такие же… наверно))))
1. Молекулярная – изуч. строение и физ-хим св-ва, биофизику молекул, биополимеров и надмол-ных систем.
2. БФ клетки – изуч. особенности строения и функц-ния кл-ных и тканевых систем. БФ мембранных процессов – св-ва био мембран и их частей; БФ фотобиол. процессов – воздействия внешних источников света на живые системы; радиационная БФ: влияния ионизирующего излучения на организм.
3. БФ сложных систем – изуч. кинетику биопроц., поведение во времени разнообразных проц-ов присущих живой материи и ТД биосистем – преобразования Е в живых структурах.
Современная БФ стремительно развивается, ее достижения способствуют переходу биологии на качественно более высокий молекулярный уровень исследования.
Не знаю, что сюда ещё, из Википедии, можно как направления: математическая БФ. Прикладная БФ: биоинформатика (хотя не является собственным разделом БФ, но очень тесно связана с ней); биометрия; биомеханика (функции и структура опорно-двигального аппарата и физ. движения биосистем); БФ эволюционных процессов; медицинская БФ; экологическая БФ.
Био объекты очень сложны и на протекающие в них процессы влияют многие факторы, кот. зависят друг от друга. Физика позволяет создать упрощенные модели объекта, кот. описываются законами ТД, электродинамики, квантовой и классической механики. С пом. корреляции физ. данных с биол-ми можно получить более глубокое понимание процессов в био объекте. Для получения инфо в био системах применяют различные оптические методы, рентгено-структурный анализ с использованием синхротронного излучения, ЯМР- и ЭПР-спектроскопию, 7-резонансную спектроскопию, различные электрометрические методы, микроэлектродную технику, методы хемилюминесценции, лазерную спектроскопию, метод меченых атомов и др. Это исп. для медицинской диагностики и терапии.
Термодинамика (ТД).
4. Термодинамика, как ядро современной биофизики. Предмет и задачи. Практическое значение ТД в БФ исследованиях.
ТД – это н. о превращения Е. ТД – это н., изуч. наиболее общие закономерности превращения различных видов Е в системе.
Предмет ТД: Е; возникновение Е в живых сист.; взаимодейств. жив. сист. с окруж. средой. Подходы: феноменологический и детальный. Значение имеют ТД параметры только в исходном и конечном состоянии.
Методы: статистический (но не даёт представление о процессе).
Направления: 1) изуч. и расчёт Е в состоянии покоя и при совершении работы. Изуч-ся и опр-ся КПД разл биол процессов. 2) изуч. динамических процессов в живых сист. (транспорт в-ва).
Значение: Позволяет оценить энергетические изменения, происх. в результате биохим. р-ций; рассчитать Е разрыва конкретных хим связей; рассчитать осмот. давление по обе стороны полупрониц. мембраны; рассчитать влияние концентрации соли в р-ре на растворимость макром-л. Применяется для описания процессов, происх. в электрохим. ячейках. Привлекается для обоснования теории возникновения и эволюции жизни на Земле.
Классификация ТД систем; особенности живых организмов, как ТД систем.
Система – любое материальное тело / объект, который сост. из большого числа частиц. Выполняют некоторую общую функцию, несводимую к ф-циям компонентов сист. Окр. среда – всё, что находится в контакте с системой.
Типы: 1) закрытые (т.е. не обмениваются в-вом): а) изолированная – сист, кот не обменивается с окр. средой не Е, не в-вом. Напр: космонавт в скафандре в открытом космосе. б) замкнутая – не обменивается с окр. средой в-вом, но обменивается Е. Напр: стакан с горячим чаем. 2) открытые – обмен и в-вом и Е. Напр: все живые сист.
Характеристика ТД функций, применяемых для анализа биолог процессов.
Функция – совокупность физ. и хим. св-в, кот. определяют конкретное сост. системы. Напр: t, V, P. ТД функции = ТД параметры. ТД параметры сост. системы – измеримые св-ва системы, связанные с Е и характеризующие её состояние. Основные параметры состояния – параметры, поддающиеся прямому измерению. Те, которые нельзя измерить непосредственно, рассматривают как функции основных параметров состояния.
Типы: 1) экстенсивные (=факторы ёмкости) – это ТД фукнции, кот зависят от кол-ва или числа частиц и их массы. Напр: масса, объём, E, энтропия (S), энтальпия (Н). Св-во системы = сумме свойств частей. 2) интенсивные – не зависят. Напр: t, давление, скорость изменения энтропии.
Состояние системы – совокупность всех физ. и хим. св-в системы.
ТД процесс – изменение сост. системы, характ-ся изм. её ТД параметров. Изменение может происходить при различных условиях, различают:
1). Равновесные и неравновесные процессы. Равн. пр. – непрерывный ряд равновесных сост. сист.; сост., не изм-ся во времени и не требующее для своего поддержания каких-либо внешних факторов. Нер. — это пр-сы, после протекания кот. систему нельзя вернуть в исходное сост. без того, чтобы в ней не осталось каких-либо изменений.
2). Обратимые и необратимые процессы. Обрю. – пр-сы, допускающие возможность возвращения системы в первоначальное сост. без того, чтобы в окр. среде остались какие-либо изменения. В противном случае – необр.
3). Самопроизвольные и несамопроизвольные процессы. Сам. – процессы, происходящие сами собой (не требующие затраты Е извне) и приближающие систему к равновесию. Несам. – процессы, требующие затраты Е извне.
Некоторые частные виды процессов:
а) изотермический (T = const);
б) изобарный (p = const);
в) изохорный (V = const);
г) адиабатический (нет обмена теплотой между системой и окр. средой);
д) изобарно – изотермический (p = const, T = const);
е) изохорно – изотермический (V = const, T = const).
ТД полного окисления глюкозы. Расчёт эффективности (КПД) биологического окисления глюкозы.
(см. вопрос 24)
При окислении глюкозы до СО2 и Н2О высвобождается больше Е, чем при гликолизе. При гликолизе: глюкоза → 2 лактат– 2Н , ∆G0 (свободная энергия) = -47,0 ккал/моль («-» т.к. выделяется). При полном окислении: глюкоза 6О2 → 6СО2 6Н2О, ∆G0= -686 ккал/моль. При гликолизе лактат содержит ≈ 93% той Е, которая была заключена в исходной м-ле глюкозы. Т.к. молочная к-та – сложное соединение, при глиголизе не происходит реального окисления. Вся био доступная Е высвобождается из глюкозы лишь в том случае, если все водородные атомы, связанные с углеродом, будут удалены и заменены кислородом с обр-нием СО2.
ЛИМИТИРУЮЩАЯ СТАДИЯ
Нередко скорость сложной химической реакции может зависеть главным образом не от всех значений констант скорости отдельных ее стадий, а только от одной. Эту стадию называют лимитирующей (т. е. определяющей скорость всей реакции в целом). Скорость последовательной реакции определяется скоростью наиболее медленной стадии, скорость же параллельных реакций определяется скоростью наиболее быстрой стадии. Например, пусть имеется сложная реакция А ® D, протекающая в три стадии:
А Р1
Р1 Р2
Р2 D.
D.
Исходное вещество А превращается в активную промежуточную частицу (АПЧ) Р1 (константа k1), которая в свою очередь превращается в АПЧ Р2 (константа k2), а уже из Р2 образуется продукт реакции — вещество D (константа k3). В том случае, если k1 << k2 » k3, то даже при малых концентрациях Р1 и Р2 скорости второй и третьей стадий очень велики и намного превышают скорость первой стадии. Фактически все образовавшееся количество Р1 очень быстро превращается в продукт реакции D. Скорость всей реакции определяется скоростью образования Р1 — лимитирующей стадией реакции будут первая:
w @ w1= k1·CA.
ЭНЕРГИЯ АКТИВАЦИИ
Зависимость скорости реакции от температуры дает уравнение Аррениуса:
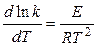 , (10.16)
, (10.16)
где k — константа скорости реакции, R — универсальная газовая постоянная, Е — энергия активации химической реакции. В случае простых реакций величина Е показывает, какой минимальной (избыточной по сравнению со средней) энергией в расчете на 1 моль должны обладать реагирующие частицы, чтобы они могли вступить в химическую реакцию. В случае сложных реакций величина Е называется эмпирической или кажущейся энергией активации и в общем случае зависит от энергий активации отдельных стадий данной реакции.
Проинтегрировав уравнение (10.16), получим уравнение Аррениуса в интегральной форме:
 , (10.17)
, (10.17)
или
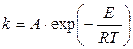 ,
,
где А — предэкспоненциальный множитель. Физический смысл А в случае простых реакций: мономолекулярных — это частота колебаний по разрываемой связи (А » 1013 сек-1), бимолекулярных — величина А пропорциональна общему числу столкновений между молекулами реагирующих веществ (А » 10-10 ¸ 10-11 см3/(мол-л·сек).
Проинтегрировав ур-е (10.16) в пределах температур от Т1 до Т2, получим:
 . (10.18)
. (10.18)
Энергию активации можно определить как аналитически по уравнению (10.18), так и графическим методом. Для этого необходимо знать ряд констант скоростей при разных температурах. Если реакция подчиняется уравнению Аррениуса, то зависимость lnk от 1/T должна выражаться прямой линией, что следует из уравнения (10.17) (рис. 7).
Рис. 7. К расчету энергии активации реакции
по температурной зависимости константы скорости.
Для очень большого числа реакций энергия активации находится в пределах от 60 до 240 кДж/моль, т.е. примерно соответствует энергиям химических связей.
Энерг актив. Связана с Q10: Е=0,46T1*T2lgQ10
ИСП УР Лайнуивера-Бэрка ДЛЯ ОПРЕДЕЛЕНИЯ КИНЕТ ХАР-ИК ФЕРМ Р-ЦИИ

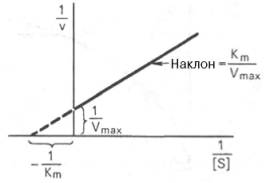 график Л-Б
график Л-Б
Графическое выражение для скорости реакции в координатах Лайнуивера-Бэрка имеет вид прямой линии, тангенс угла наклона который будет равен величине Km/Vmax . Линия отсек на оси Х значение -1/Km, а на оси Y- значение 1/V max
АНАЛ НЕКОНКУР ИНГИБИР
НЕКОНКУРЕНТНОЕ ИНГИБИРОВАНИЕ ПРОЯВЛЯЕТСЯ ПРИ СВЯЗЫВАНИИ ИНГИБИТОРА С ФЕРМЕНТОМ ВНЕ АКТИВНОГО ЦЕНТРА, но при этом меняется структура активного центра и связь с субстратом становится невозможной.
E I —> EI, EI S —-> (невозможно)
 скор р-ции
скор р-ции
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси абсцисс. ингибитор не изменяет Кm и снижает V max.

Ренатурация (renaturation)
– Восстановление (после денатурации) нативной (биологически активной) пространственной структуры биополимера (белка или нуклеиновой кислоты). В частности, ренатурация ДНК (после денатурации нагреванием) может происходить при медленном охлаждении, что используется для получения гибридных гетеродуплексов.
Метод дифференциальной диагностики и прямого выявления нуклеиновых кислот в исследуемом материале с помощью реакции молекулярной гибридизации и полимеразной цепной реакции (ПЦР).
Диагностическая суть этих методов заключается в том, что у исследователей имеются определенные олигонуклеотиды, воспроизводящие (или комплементарные), высококонсервативные и высокоспецифичные последовательности нуклеотидовиз искомого генома. В реакции молекулярной гибридизации такие олигонуклеотилы называют зондами, в методе ПЦР -праймерами. В реакции молекулярной гибридизации назначение этих специфических олигонуклеотидов-“найти” в исследуемой нуклеиновой кислоте комплементарные последовательности, и сформировать структуры согласно природе комплементарных взаимодействий между парами азотистых оснований . Материалом для исследований служит кровь, культуральная жидкость, ликвор, биопсийный материал, носоглоточные смывы, моча и пр. биологические субстанции. Молекулярно-генетические методы перспективны при необходимости ранней диагностики ВИЧ до появления в сыворотке AT, при неясных серологических результатах, при определении наличия ВИЧ в организме новорожденных детей от инфицированных женщин. Целесообразно использовать эти методы при обследовании серонегативных сексуальных партнеров инфицированных лиц
Электрокинетический потенциал: возникновение, измерение и факторы, определяющие его величину. Примеры электрокинетических явлений, их характеристика и научно-практическое значение.
Потенцил Действия
1. Объясняется поворотом диполя на 1800.
2. Теория альтерации Германа. При возбуждении возникает избыток кислых продуктов, которые несут отрицательны заряд, что приводит к разности потенциалов между возбужденным и невозбужденым учаском.
3. Мембранная теория Берштейна. В возбужденном участке мембраны резко увеличивается проницаемость для всех ионов, концентрации ионов смешиваются и участок становится электронейтральным.
4. Ходжкин и Хаксли. Рост проницаемости мембраны для ионов в месте воздействия. При возбуждении электропроницаемость мембраны увеличивается примерно в 500 раз. Max увеличивается проницаемость мембраны для Na (отсюда Na-теория ПД). Na свободно проходит внуть клетки. При возбужедении электро-химическое равновесие определяется потенциалом Na . Равновесный потенциал для К = –97 мВ, для Na = 50 мВ. При возбужедении мембрана перезаряжается. Положение обратной активации и инактивации Na -каналов, Na -канал может активироваться (открываться) при определенных значениях потенциала. Причина активации Na -каналов – деполяризация мембраны, чем больше деполяризация, тем больше проницаемость мембраны для Na . Зависимость близка к линейной в подкор уровне; как только мембрана достигнет критического уровня деполяризации – зависимость нелинейная, лавинообразный вход Na в клетку.
Электрокинетический потенциал, x-потенциал, дзета-потенциал, часть общего скачка потенциала на границе двух фаз, определяющая относительное перемещение этих фаз при электрокинетических явлениях. Общий скачок потенциала при пересечении межфазной границы в дисперсных системах обусловлен существованием двойного электрического слоя. Электрокинетический потенциал — перепад потенциала по той части диффузного слоя, в пределах которой жидкость может быть вовлечена в тангенциальное движение относительно межфазной поверхности при внешнем воздействии на систему. Под влиянием сильно адсорбирующихся на поверхности ионов или изменения pH жидкости может произойти перемена знака на противоположный («перезарядка» поверхности). Электрокинетический потенциал в изоэлектрической точке равен нулю. он является одной из важнейших характеристик жидких коллоидных систем. Его значение и характер изменения при варьировании параметров электролита, адсорбции на пов-сти разл. в-в и т. п. позволяет судить о структуре граничных слоев, особенностях взаимод. компонентов р-ра с пов-стью, заряде пов-сти и т.д.
1). Для объяснения реполяризации используется положение об инактивации Na -каналов. При приближении потенциала мембраны к равновесному для Na , Na -каналы инактивируются и посупление Na в клетку прекращается. К графику: в основе регенеративный процесс (сам себя поддерживающий), развивающийся по принципу обратной связи.
2). Рост К проницаемости мембраны. Не столь значителен, как для Na ( в 5-15 и 500 раз соответственно). Проницаемость для К развивается медленнее, чем для Na . Ионы К в этой ситуации будут выходить наружу и выносить заряд.
3). Механизм активного транспорта, представленный K-Na-насосом. 3 Na внутрь и 2 К наружу.
Эксперименты Ходжкина и Хаксли.
Гигантский аксон кальмара. Из внеклеточной среды были удалены 2/3 Na . При этом амплитуда ПД снизилась ? на 50%. Замена внутриклеточного Na на другие ионы приводит к некоторому росту ПД. Замена ? внутриклеточного К на Na приводит к значительному снижению ПД.
Метод фиксации потенциала
метод Петч-Клемпинга. С его помощью можно зафиксировать на длительное время значение мембранного потенциала на любом желаемом уровне. Это делается с помощью внешнего генератора напряжения
Суммарные мембранные токи при ПД
1. Подпороговая область:
Слабое изменение мембранного потенциала, суммарный ионный ток направлен от клетки наружу, так как поток К , выходящий из клетки, уже усиливается из-за удаления мембранного потенциала от равновесного потенциала для К . Входящий ток Na еще слаб, так как рост Na -проницаемости пока невелик. Однако с развитием деполяризации Na-ый поток постепенно нарастает.
2. Критический уровень деполяризации:
В этот момент суммарный ионный ток через мембрану равен нулю, так как встречные токи ионов Na и К уравновешивают друг друга. Даже небольшая дальнейшая деполяризация приводит к росту входа Na -тока в сотни раз.
3. Во время фазы деполяризации резко увеличивается Na -проницаемость и суммарный мембранный ток, направленный внутрь клетки. Выходящий К -ток растет медленнее и становится заметным только к моменту пика потенциала.
4. Фаза реполяризации:
В момент пика потенциала большинство Na -каналов инактивированны, а К -ток max. Поэтому суммарный мембранный ток – выходящий.
Кальциевая теория активации и инактивации Na -каналов
В состоянии покоя у наружного отверстия Na -канала находится Са2 , который электростатически тормозит проникновение Na в канал. При возбуждении наружная поверхность мембраны заряжена отрицательно, при этом Са2 уходят со своих мест, вход открывается и Na входит в клетки.
Инактивация: по ходу деполяризации узкие Na -каналы могут закупориваться Na . Во многих каналах есть воротные белки (могут менять свое местоположение под влиянием изменения потенциала). В состоянии покоя активационный белок закрыт, а инактивационный открыт. При возбуждении открывается активационный белок в момент закрывания инактивационного белка. В конце реполяризации белки так же закрываются и потом открываются (исходное состояние).
Факторы, определяющие скорость распространения возбуждения по нервному волокну
1. Пространственная константа определяет величину декремент
деполяризации, ? – пространственная константа.
2. Коэффициент надежности, соотношение между амплитудой ПД
критической энергией и ПП. S=ПД/(Екр–ПП), ПД=120мВ, ПП=–70мВ, Екр=–55мВ ? S=8. Чем больше S, тем быстрее проведение.
3. Временная константа ? мембр. При возбуждении мембраны меняется
заряд. Длительность перезарядки мембраны. ?мембр=Rm*Cm. Чем больше ? мембр, тем ниже С. Vраспр=S*?/?мембр.
Биологические и физические процессы и закономерности в живых системах. Редукционизм и антиредукцианизм. Принцип качественной несводимости.
Живые огранизмы – открытая, саморег-ся, самовоспр-ся и разв-ся гетерогенная система, важнейшими функциональными в-вами в которой явл-ся биополимеры: белки и нукл. к-ты сложного атомно-мол-ного строения.
Первые попытки объяснения биолог. пр-сов связаны с методами сравнения с физич. пр-ми. Напр: м-м нервного проведения – как распростр. волны окисления в медной проволоки в кислоте; мышечное сокращение объяснялось работой пьезо элементов; рост клетки. Первоначально физика проникла в химию – необходимость объяснить как разл. соединения взаимод-ют в организме – физическая химия и химическая физика.
Сущ 2 лагеря современ. физики:
1) Редукционисты: Любой био процесс происх. в жив организме можно свести к суме хим., физ. и механич. процессов. Объяснение сложного ч/з более простое, непонятное ч/з известное. Зная св-ва отдельных элементов, сост. систему и особенности их взаимод-вия, можно вывести все свойства этой системы. Обр. более сложного уровня – это результат усложнения более простых. Иногда: попытки заменить исследование реального объекта его упрощенной моделью. Достижение: предсказание существования планеты Нептун. Но как метод мышления не является универсальным. Неудача в биологии: не могут объяснить с этой т.зр. феномен жизни.
2) Антиредукционисты: Принцип качественной несводимости или био антиред., т.е. невозможность свести законы и принципы, управляющие живой материей к элементарной сумме физико-хим и мех. процессов процесс жизнедеятельности. Т.е. физико-матем. модели не м.б. адекватными, если в них не сод-ся элементы функциональной организации живых систем. Т.е. существует предел, после которого физические представления перестают быть самодостаточным средством познания, а дальше определяющим фактором становятся некоторые био истины, без которых уже не обойтись.
§
( см вопрос 17)
Стационарное сотояние:
Зависит от 3-х параметров: -S (энтропия), -U (своб. E), -t (время).
Стац. сост. – сост., когда параметры системы (U, S) с теч. времени остаются неизменными, но происходит обмен в-вом и Е, т.е. сост. сист. при кот. не происх. изм. ТД параметров.
Изолированные сист.: ∆S=0 или >0.
Для равновесного сост. S стремится к мах, U=0. Стац. сост. отличается тем, что S ≠ мах, а является постоянной величиной, S=const, U не равняется 0, U=const. Ежесекундный прирост энтропии стремится к min. Любая живая система может находиться только в стац. сост. Если достигнуто состояние ТД равновесия – это уже не живая система. Качество стационарного состояния может быть различным.
В открытых системах:
S состоит из двух показателей.
Si – внутри самой сист., S – самой системы, Se – внешняя среда.
dS=dSi dSe (d – это ∆ – это изменение)
Когда dSe > dSi и dSe < 0, тогда dSe < 0 – «нек» энтропия, негативная, в эволюц. плане деградирует система, напр. паразиты.
В случае изомерной живой системы – она не обменивается Е и в-вом, т.е. только dS=dSe
Прирост S – необратимость протекания процесса.
dSe<0: возможно 3 ситуации
1.Приток внешней энтропии отрицателен и по модулю превышает изменения внутренней Si dS<0
нервный импульс.
2. dSe<0, по мoдyлю=dSi
dS=O
Характерно для стационарного состояния системы dS=dSi dSe=O
3. dSe<0 и по мoдyлю <dSi. dS>0.
Состоянию ТД равновесия – характерно мах значение S (S=max), U=0, т.е. Е, которая расходуется на совершение А.
Сходство: стац. и равновесное состояния не зависят от времени.
Отличия стац. сост. от равновесия (из конспекта):
1) своб. Е (∆G) в стац. сост. есть величина постоянная во времени и не равна 0. В ТД равн. ∆G=const, но ∆G =0 => открытые сист., если вывести из стац. сост. могут совершать работу; при ТД равновесии не способны совершать работу.
2) энтропия. В стац. сост. =const, но она не max. (∆G) ∆S ≠ max = const.
3) !!! в стац. сост. проявляется кинетический параметр (фактор) (изменение энтропии во времени) dS/dt = dSi/dt dSe/dt.
Стационарное состояние:
* постоянный обмен энергией с окружающей средой
* постоянно тратится свободная энергия на поддержание состояния
* т/д потенциалы постоянны, G и F не равны 0
* энтропия постоянна, но не максимальна
* градиенты присутствуют
Термодинамическое равновесие
* отсутствует поток вещества и энергии в окружающую среду и обратно
* на поддержание этого состояния не затрачивается свободная энергия
* работа способности системы равна 0, т/д потенциалы равны 0
* энтропия максимальна
* в системе отсутствуют градиенты
Переход на новый стац. уровень:
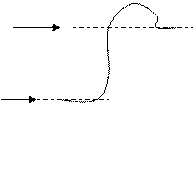 2 пути: 1) «овершот» – по нему переходят живые организмы при изм внеш. усл. (приспособление). График.
2 пути: 1) «овершот» – по нему переходят живые организмы при изм внеш. усл. (приспособление). График.
Нижняя стрелочка – это старый стац. уровень.
Верхняя стрелочка – это новый стац. уровень.
2) «ложный старт» – усиление или уменьшение О2, выращивание лука с О2 и без. График. С О2 – аэробный распад углеродов. Без О2 – обмен в-в переходит на анаэробный путь. А если потом снова дать О2 – то получится график 2 (то что обведено кружочком – там осущ-ся уничтожение продуктов анаэробного пути). Пример для чела: пока не расщепится молочная к-та осуществлять работу дальше нельзя.
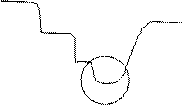

17. Теорема Пригожина и направленность эволюции биосистем. Энтропия и биологический прогресс.
( см вопрос 16)
Стац. сост. хар-ся min ежесекундным приростом энтропии (благодаря этому происходит эволюция).
Теорема: при постоянных внеш. усл. в системе, находящейся вблизи положения ТД равновесия в стац. сост., скорость возрастания энтропии, за счёт необходимости внутр. процессов, принимает постоянное минимальное значение отличное от нуля.
Или: В стационарных состояниях при фиксированных внешних параметрах локальная продукция энтропии в открытой т/д системе стремится к минимальному значению.
Энтропия – мера рассеивания свободной энергии, следовательно любая открытая т/д система в стационарном состоянии стремится к минимальному рассеиванию свободной энергии. Если в силу причин система отклонилась от стационарного состояния, то вследствие стремления системы к минимальной энтропии, в ней возникают внутренние изменения, возвращающие ее в стационарное состояние.
Величина, кот это всё характеризует:
β= T* (dS/dt), где β – диссипативная фукнкция. β>0, min. С этим связан Критерий эволюции открытых систем: ∆β/dt < 0
Механизмы саморегуляции систем
Функционируют по принципу обратной связи. Обратная связь – это понятие, обозначающее влияние выходного сигнала системы на ее рабочие параметры. Различают положительную и отрицательную. “–” чаще встречается в биосистемах, направлена на снижение влияния выходного сигнала на рабочие параметры системы. ” ” усиливает влияние выходного сигнала, в результате чего система может выходить из данного состояния.
Гомеостаз – постоянство многих параметров.
§
Гликолиз – это серия реакций, в результате которых глюкоза распадается на две молекулы пирувата (аэробный гликолиз) или две молекулы лактата (анаэробный гликолиз).
!!! Пример ферментативной р-ции: Фермент (E) субстрат (S) → ES → P(уходит) E (фермент остаётся). Относится к циклическим пр-сам, выгодно, т.к. затратно каждый раз делать новые ферменты, малодоступны, витамины.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание О2 недостаточно, напр. в активно сокр-ся мышце, пируват превращается в лактат.
Конечным продуктом гликолиза является молочная кислота. Суммарное уравнение:
C6H12O6 2 АДФ 2 Фн → 2СН3СН(ОН)СООН (молочная к-та) 2АТФ 2 Н2О.
Это уравнению можно разбить на 2: 1) глюкоза → 2 лактат– 2Н , ∆G1 (свободная энергия) = -47,0 ккал/моль («-» т.к. выделяется); 2) 2 АДФ 2 Фн → 2АТФ 2 Н2О, ∆G2 = 2 7,3= 14,6 ккал/моль (т.к. требует затрат Е). Эти процессы сопряжены. Превращение 1 моль глюкозы в лактат в станд. усл. приводит к высвобождению гораздо большего кол-ва св. Е (47,0 ккал), чем необх. для обр. АТФ из АДФ (14,6 ккал). В живой кл. при истинных внутрикл. концентрациях АТФ, АДФ и Фн, глюкозы и лактата эффективность запасания высвобождающейся при гликолизе Е превышает 60%.
Определим общее изменение станд. свободной Е: ∆Gs= ∆G1 ∆G2 = -47 14.6 = -32.4 ккал/моль. => суммарная р-ция гликолиза сопровождается очень большим снижением свободной Е.
При гликолизе лактат содержит ≈ 93% той Е, которая была заключена в исходной м-ле глюкозы. Т.к. молочная к-та – сложное соединение, при глиголизе не происходит реального окисления.
Благодаря гликолизу организм чела и животных в определен. период может осуществлять ряд физиолог. функций в условиях недостаточности кислорода.
КПД = 40% (36%), 80 кДж.
Последовательность реакций хорошо изучена. Процесс гликолиза катализируется одиннадцатью ферментами. Протекает в гиалоплазме (цитозоле) клетки. Все 11 р-ций ферментативные.

Био значение процесса гликолиза: образовании богатых Е фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 м-лы АТФ (гексокиназная и фосфофрук-токиназная р-ции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная р-ции). Энергетическая эффективность гликолиза в анаэробных условиях составляет 2 м-лы АТФ на одну м-лу глюкозы.
Основной р-цией, лимитирующей скорость гликолиза, является фосфофруктокиназная. Затем – гексокиназная р-ция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами.
ТД характеристика окисления пировиноградной кислоты в цикле Кребса. Расчёт КПД.
Цикл трикарбоновых кислот открыт англ. биохимиком Г. Кребсом. Значение данного цикла: полного сгорания пирувата, главным источником кот. является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбоновых кислот является тем центром, в котором сходятся практически все метаболические пути. Цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть орг. м-л, играющих роль «клеточного топлива»:углеводов, жирных кислот и аминокислот.
!!! Пример ферментативной р-ции: Фермент (E) субстрат (S) → ES → P(уходит) E (фермент остаётся). Относится к циклическим пр-сам, выгодно, т.к. затратно каждый раз делать новые ферменты, малодоступны, витамины.
ПВК (образовался при гидролитическом расщеплении глюкозы) окисляется до ацетил – СоА и СО2 при участии ферментов (они наз-ся пируватдегидрогеназный комплекс). Происходит окислительное декарбоксилирование ПВК. ∆G0 (свободная энергия)= -8 ккал/моль («-» т.к. выделяется).
Если в пpоцессе гликолиза (по данным A.L. Lehninger, 1972) пpи pаспаде 1 моля глюкозы высвобождается 50 ккал, или 208 кДж свободной энеpгии, то пpи окислительном pаспаде 1 моля глюкозы в ЦТК – около 689 ккал, или 2883 кДж свободной энеpгии.
Образуется 12 молекул АТФ при сжигании 1 молекулы ацетильного радикала.
Скорость протекания хим. процессов определяется уровнем температуры.
Цикл происходит в матриксе митохондрий и состоит из 8 последовательных реакций. В результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула оксалоацетата регенерируется.

Весь оборот цикла Кребса приводит к исчезновению одной молекулы ПВК, причем углерод и кислород выделяются в виде углекислого газа, а отщепляемый от органического субстрата водород подвергается дальнейшим превращениям, и в конечном счете, будучи энергетически обесцененным, соединяется с молекулярным кислородом, образуя воду.
ТД полного окисления глюкозы. Расчёт эффективности (КПД) биологического окисления глюкозы.
(см. вопрос 24)
При окислении глюкозы до СО2 и Н2О высвобождается больше Е, чем при гликолизе. При гликолизе: глюкоза → 2 лактат– 2Н , ∆G0 (свободная энергия) = -47,0 ккал/моль («-» т.к. выделяется). При полном окислении: глюкоза 6О2 → 6СО2 6Н2О, ∆G0= -686 ккал/моль. При гликолизе лактат содержит ≈ 93% той Е, которая была заключена в исходной м-ле глюкозы. Т.к. молочная к-та – сложное соединение, при глиголизе не происходит реального окисления. Вся био доступная Е высвобождается из глюкозы лишь в том случае, если все водородные атомы, связанные с углеродом, будут удалены и заменены кислородом с обр-нием СО2.
§
В отличие от простых (одностадийных) реакций, уравнение которых совпадает со стехиометрическим уравнением реакции, подавляющее большинство химических реакций является сложными, т.е. протекает через ряд последовательных и (или) параллельных стадий, при этом некоторые стадии протекают с участием промежуточных веществ, не являющихся ни исходными веществами, ни продуктами реакции.
В основе кинетики сложных реакций лежит принцип независимости реакций, согласно которому “если в системе одновременно протекает несколько реакций, то каждая из них независима от остальных и скорость ее прямо пропорциональна произведению концентраций реагирующих веществ”. Этот принцип справедлив для обратимых, параллельных и последовательных реакций. Конечное изменение концентрации данного вещества является результатом всех независимых изменений.
Однако принцип независимости реакций не является таким общим как основной постулат химической кинетики. Он не применим, например, к сопряженным реакциям, в которых одни простые реакции существенно влияют (часто ускоряют ) на протекание других реакций.
Все процессы живых систем многостадийные:
3 группы
1. последовательные реакции: 1 исходное вещество которое претерпевает превращения (глюкоза до молоч.кислоты, молоч.кислота до ПВК)
Последовательные реакции протекают через ряд стадий, в каждой из которых претерпевает превращение продукт, образовавшийся в предыдущей стадии. К последовательным реакциям относят реакции типа:
АВD.
Вещество В образуется из А (константа k1) и расходуется на образование D (константа k2), обе реакции мономолекулярны. Скорость изменения концентрации вещества В со временем будет равна:
 ,
,
где С0А — начальная концентрация вещества А (начальные концентрации В и С равны нулю); х и y — число молей веществ А и В, соответственно, прореагировавших к моменту времени t.
2. параллел.р-ции:
Параллельными называют реакции, в которых одни и те же исходные вещества реагируют одновременно в двух или более направлениях. Это реакции типа
В АС.
АС.
Исходное вещество А одновременно расходуется на образование вещества В (константа k1) и вещества С (константа скорости k2). Обе реакции первого порядка. Суммарная скорость расходования вещества А равна
 , (10.20)
, (10.20)
где w1 и w2 — скорости превращения вещества А в вещества В и С, соответственно; С0А — начальная концентрация А; х — количество вещества А, прореагировавшего к моменту времени t.
После интегрирования ур-я (10.20) получаем
 . (10.21)
. (10.21)
По уравнению (10.21) можно рассчитать сумму констант (k1 k2). Чтобы разделить эту сумму на отдельные составляющие, необходимо использовать еще одно уравнение, дающее отношение констант в некоторый момент времени t:
 , (10.22)
, (10.22)
где х1 и х2 — числа молей веществ В и С, образовавшихся к моменту времени t (х = х1 х2). Решая систему уравнений (10.21) и (10.22), можно найти значение каждой константы скорости по отдельности.
ЛИМИТИРУЮЩАЯ СТАДИЯ
Нередко скорость сложной химической реакции может зависеть главным образом не от всех значений констант скорости отдельных ее стадий, а только от одной. Эту стадию называют лимитирующей (т. е. определяющей скорость всей реакции в целом). Скорость последовательной реакции определяется скоростью наиболее медленной стадии, скорость же параллельных реакций определяется скоростью наиболее быстрой стадии. Например, пусть имеется сложная реакция А ® D, протекающая в три стадии:
АР1Р2D.
Исходное вещество А превращается в активную промежуточную частицу (АПЧ) Р1 (константа k1), которая в свою очередь превращается в АПЧ Р2 (константа k2), а уже из Р2 образуется продукт реакции — вещество D (константа k3). В том случае, если k1 << k2 » k3, то даже при малых концентрациях Р1 и Р2 скорости второй и третьей стадий очень велики и намного превышают скорость первой стадии. Фактически все образовавшееся количество Р1 очень быстро превращается в продукт реакции D. Скорость всей реакции определяется скоростью образования Р1 — лимитирующей стадией реакции будут первая:
w @ w1= k1·CA.
§
Предварительные эксперименты по изучению кинетики ферментативных реакций показали, что скорость реакции E S —> E P, вопреки теоретическим ожиданиям, не зависит от концентрации фермента и субстрата так, как в случае обычной реакции второго порядка. Самая ранняя попытка математически описать ферментативные реакции была предпринята Дюкло в 1898 г. Браун (1902) и независимо от него Анри (1903) впервые выдвинули гипотезу об образовании в ходе реакции фермент-субстратного комплекса. Это предположение основывалось на трех экспериментальных фактах:
1. папаин образовывал нерастворимое соединение с фибрином
2. субстрат инвертазы – сахароза могла защищать фермент от тепловой денатурации
3. было показано, что ферменты являются стереохимически специфическими катализаторами
Х-КА ДЕЙСТВИЯ ФЕРМ:
1. специфичность действия-способность ускорять протекание 1 или нескольких реакций (амилазная реакция расщепляет крахмал до глюкозы)
а. абсолютная- определенный субстрат;
б. относительная- ферменты, которые катализируют ращепление определенного типа связи. (пепсин)
2. ускорение протекания ферментативн. Реакций- каталитичность. Ферменты действуют в мыгких условиях (норм давление, pH, температура): гидролиз крахмала
Амилаза = 37 градусов, pH7, скорость выше, чем при неорган.катализе.
2. регулируемость – есть факторы под воздействием которых скорость может увеличится или уменшаться
В 1913 году Михаэлис и Ментен опубликовали свою теорию общего механизма ферментативных реакций. Их уравнение стало фундаментальным принципом всех кинетических исследований ферментов вот уже почти целый век.

т.е. фермент Е вступает во взаимодействие с субстратом S с образованием промежуточного комплекса ES, который далее распадается на свободный фермент и продукт реакции Р. Математическая обработка на основе закона действующих масс дала возможность вывести уравнение, названное в честь авторов уравнением Михаэлиса–Ментен, выражающее количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции:

где v – наблюдаемая скорость реакции при данной концентрации субстрата [S]; KS– константа диссоциации фермент-субстратного комплекса, моль/л; Vmax– максимальная скорость реакции при полном насыщении фермента субстратом.
Основной механизм действия ферментов-они снижают Е активации за счет образования фермент-субстрат копмлекса.
Катализ приводит к ускорению достижения равновесия за счет снижения энергии активации (Еа), часто ступенчато.
ВЫВОД И АНАЛИЗ УР МИХАЭЛИСА-МЕНТЕН
Михаэлис и Ментен предположили, что мех-м ферм р-й описывается моделью:
 (2.1)
(2.1)
При формулировке кинетического выражения для скорости ферментативной реакции Михаэлис и Ментен сделали три допущения:
1) Стационарное состояние реакции в момент равновесия, когда скорости образования и расходования ES равны;
2) Весь фермент в условиях насыщающих концентраций субстрата превращается в энзимсубстратный комплекс ES;
3) Если весь фермент в виде ES, то скорость реакции максимальна и Vmax=k2[ES].
Образование ES: [ES]=k1[S][E] (I)
Расходование ES: [ES]=k-1[ES] k2[ES] (II)
Приравнивая выражения (I) и (II) и сокращая обе части на k1 получаем:
[S][E] = [ES](k-1 k2)/k1 = [ES]Km, где Km = (k-1 k2)/k1
Выразим равновесную концентрацию [E] через начальную [Eo]: [E] = [Eo] – [ES]
[S]([Eo]-[ES])= [ES]Km, переносим [S] в правую часть выражения и делим обе части на [ES]:
[Eo]/[ES]=Km/[S] 1= (Km [S])/[S] (III)
Поскольку трудно (если не невозможно) измерить [ES], произведем замену с учетом того, что в насыщающих концентрациях [S] весь [Eo] перейдет в [ES] и максимальная скорость при этом будет равна Vmax=k2[ES]=k2[Eo].
В это же время скорость реакции равна V=k2[ES]. Через отношение этих скоростей выразим [Eo]/[ES]:
V/Vmax= [ES]/[Eo]
В уравнении (III) произведем замену отношения [Eo]/[ES] на Vmax/V и получаем:
V = Vmax[S]/(Km [S])
Это и есть уравнение Михаэлиса-Ментен.
§
Осмотическое давление, диффузное давление, термодинамический параметр, характеризующий стремление раствора к понижению концентрации при соприкосновении с чистым растворителем вследствие встречной диффузии молекул растворённого вещества и растворителя. Если раствор отделен от чистого растворителя полупроницаемой мембраной, то возможна лишь односторонняя диффузия — осмотическое всасывание растворителя через мембрану в раствор. В этом случае О. д. становится доступной для прямого измерения величиной, равной избыточному давлению, приложенному со стороны раствора при осмотическом равновесии. О. д. обусловлено понижением химического потенциала растворителя в присутствии растворённого вещества. Тенденция системы выравнивать химические потенциалы во всех частях своего объёма и перейти в состояние с более низким уровнем свободной энергии вызывает осмотическое (диффузионный) перенос вещества. О. д. в идеальных и предельно разбавленных растворах не зависит от природы растворителя и растворённых веществ; при постоянной температуре оно определяется только числом «кинетических элементов» — ионов, молекул, ассоциатов или коллоидных частиц — в единице объёма раствора. Первые измерения О. д. произвёл В. Пфеффер (1877), исследуя водные растворы тростникового сахара. Его данные позволили Я. X. Вант-Гоффу установить (1887) зависимость О. д. от концентрации растворённого вещества, совпадающую по форме с Бойля — Мариотта законом для идеальных газов. Оказалось, что О. д. (p) численно равно давлению, которое оказало бы растворённое вещество, если бы оно при данной температуре находилось в состоянии идеального газа и занимало объём, равный объёму раствора. Для весьма разбавленных растворов недиссоциирующих веществ найденная закономерность с достаточной точностью описывается уравнением: pV = nRT, где n — число молей растворённого вещества в объёме раствора V; R — универсальная газовая постоянная; Т — абсолютная температура. В случае диссоциации вещества в растворе на ионы в правую часть уравнения вводится множитель i > 1, коэффициент Вант-Гоффа; при ассоциации растворённого вещества i < 1. О. д. реального раствора (p’) всегда выше, чем идеального (p”), причём отношение p’/ p” = g, называемое осмотическим коэффициентом, увеличивается с ростом концентрации. Растворы с одинаковым О. д. называется изотоническими или изоосмотическими. Так, различные кровезаменители и физиологические растворы изотоничны относительно внутренних жидкостей организма. Если один раствор в сравнении с другим имеет более высокое О. д., его называют гипертоническим, а имеющий более низкое О. д. — гипотоническим.
О. д. в клетках животных, растений, микроорганизмов и в биологических жидкостях зависит от концентрации веществ, растворённых в их жидких средах. Солевой состав биологических жидкостей и клеток, характерный для организмов каждого вида, поддерживается избирательной проницаемостью биологических мембран для разных солей и активным транспортом ионов. Относительное постоянство О. д. обеспечивается водно-солевым обменом, т. е. всасыванием, распределением, потреблением и выделением воды и солей (см. Выделение, Выделительная система, Осморегуляция). У т. н. гиперосмотических организмов внутреннего О. д. больше внешнего, у гипоосмотических — меньше внешнего; у изоосмотических (пойкилоосмотических) внутреннее О. д. равно внешнему. В первом случае ноны активно поглощаются организмом и задерживаются в нём, а вода поступает через биологич. мембраны пассивно, в соответствии с осмотическим градиентом. Гиперосмотическая регуляция свойственна пресноводным организмам, мор. хрящевым рыбам (акулы, скаты) и всем растениям. У организмов с гипоосмотической регуляцией имеются приспособления для активного выделения солей. У костистых рыб преобладающие в океанических водах ионы Na и Cl— выделяются через жабры, у морских пресмыкающихся (змеи и черепахи) и у птиц — через особые солевые железы, расположенные в области головы. Ионы Mg2 ,  , у этих организмов выделяются через почки. О. д. у гипер- и гипоосмотических организмов может создаваться как за счёт ионов, преобладающих во внешней среде, так и продуктов обмена. Например, у акуловых рыб и скатов О. д. на 60% создаётся за счёт мочевины и триметиламмония; в плазме крови млекопитающих — главным образом за счёт ионов Na и Cl—; в личинках насекомых — за счёт разнообразных низкомолекулярных метаболитов. У морских одноклеточных, иглокожих, головоногих моллюсков, миксин и др. изоосмотических организмов, у которых О. д. определяется О. д. внешней среды и равно ему, механизмы осморегуляции отсутствуют (исключая клеточные).
, у этих организмов выделяются через почки. О. д. у гипер- и гипоосмотических организмов может создаваться как за счёт ионов, преобладающих во внешней среде, так и продуктов обмена. Например, у акуловых рыб и скатов О. д. на 60% создаётся за счёт мочевины и триметиламмония; в плазме крови млекопитающих — главным образом за счёт ионов Na и Cl—; в личинках насекомых — за счёт разнообразных низкомолекулярных метаболитов. У морских одноклеточных, иглокожих, головоногих моллюсков, миксин и др. изоосмотических организмов, у которых О. д. определяется О. д. внешней среды и равно ему, механизмы осморегуляции отсутствуют (исключая клеточные).
Диапазон средних величин О. д. в клетках организмов, не способных поддерживать осмотический гомеостаз, довольно широк и зависит от вида и возраста организма, типа клеток и О. д. окружающей среды.
§
Потенциал покоя
это разница потенциалов между цитоплазмой и внеклеточной средой, существующая в каждой живой клетке, находящейся в состоянии покоя. Внутриклеточная среда заряжена отрицательно. Величина ПП может быть различной, зависит от состояния клетки от –15 до –90 мВ у большинства клеток.
Для того,чтобы измерить величину ПП используется микроэлектродная техника. Используется специальный вид электродов, отличающийся намного меньшим диаметром кончика (доли мкм, или 1 мкм). Бывают стеклянные и металлические, по форме напоминают копье, необходим раствор электролита для хорошего проведения электрического тока.
Основные теории ПП:
1. В 1848 г. Дюбуа-Реймон “теория электромоторных молекул” (теория заряженных диполей). Дюбуа-Реймон предположил, что на мембране находятся диполи, ориентированные отрицательным зарядом внутрь. При возбуждении диполи поворачиваются на 1800, что приводит к положительному заряду внутри клетки. Разность потенциалов в этой теории является предшествующей.
2. В 60-е годы XIX в. – альтернативная теория Германа. В состоянии покоя
мембрана клетки не заряжена, и ПП отсутствует. Однако при повреждении, в поврежденном участке появляется избыток отрицательного заряда, в силу наличия кислых продуктов. Поэтому между поврежденными и неповреждеными участками возникает электрический ток. Теория Германа исключала существование разности потенциалов на мембране.
3. Теория полупроницаемой мембраны Берштейна. 1906 г. Положения:
~ мембрана обладает свойством полупроницаемости (в специальных экспериментах в группе Пфейффера показали: при пропускании электрического тока – поляризация мембраны и появление концентрационной электро-двигательной силы.
~ наличие концентрационных градиентов на биологической мембране. В исходном состоянии К , Сl–, Na и другие распределены различно на мембране. Берштейн обратил внимание на К -концентрационный градиент по направлению из клетки наружу.
Существуют механические насосы. Перемещение из клетки через мембрану осуществляется по градиенту; но выход не бесконечен, как только К выйдет из клетки, образуются силы, противодействующие этому выходу. 1. Первый положительный заряд на мембране препятствует выходу остальных (электростатическое отталкивание). 2. Крупные молекулы, которые не могут проходить через мембрану, мембраны их не пропускают, они не будут пропускать К мембранный слой ионов К снаружи, слой анионов изнутри. При повреждении мембраны анионы выходят наружу и создается электрический ток.
Еm = RT/F * ln([K ]ant/[K ]in)
Еm – ПП, F- число Фарадея.
Передача возбуждения по нервным волокнам
В начале 30х годов ХХ в. Хилл. 1932 г. “Химическая волна проведения в нервах”. Хилл использовал разные нервы, но преимущественно краба. Даже в состоянии покоя в единицу времени вырабатывается некоторое количество тепла. Это тепло было названо теплопродукция покоя. Когда в нервном волокне возбуждение – теплопродукция возбуждения (ТВ), она делится на 2 фазы:
1. Начальная ТВ, которая составляет 2-3% от всей ТВ и приходится непосредственно на период возбуждения.
2. Задержанная ТВ ? 97% всей ТВ. Если подать серийный импульс на нерв краба, то задержанную ТВ можно зафиксировать в течение 25-30 минут. Возбуждения в тканях уже нет, но ТВ имеет место.
3. Утечка тепла при работе Na.
Хилл разрабтал чувствительную теплоэлектрическую методику, которая позволяла фиксировать теплообразование в течение 20 мс. Эксперименты при О0 С. Начальную фазу теплопродукции делили на 2 периода: позитивная и негативная начальная теплопродукция. При О0 С для нерва краба позитив в начальные 20 мс = 14 мк кал. В течение последующих 150 мс ? 85% тепла поглощается нервной тканью обратно (12 мк кал).
Позитивная начальная теплопродукция: причина: химические процессы, обуславливающие изменение проницаемости мембраны. При возбуждении в клетку поступает Na и смешивается с К и наоборот. Должно образовываться тепло. Это тепло покрывает до 50% позитивной начальной теплопродукции.
Негативная начальная теплопродукция: химические реакции в этот период могут быть эндотермическими. Негативная теплопродукция не является обязательной.
Проведение возбуждения
В 1885 г. Герман предложил теорию малых токов. Осуществляется последовательно между участками волокна. В участке, соседнем с возбужденным будет наблюдаться выход электрического тока.
Кабельная теория нервного волокна: нервное волокно внутри содержит проводящую среду, оболочка невного волокна имеет слой, который плохо проводит возбуждение. Нервное волокно омывается внеклеточной жидкостью, которая проводит электрический ток.
Эквивалентная электрическая схема нервного волокна
В состоянии покоя внутриклеточная среда имеет избыточный отрицательный заряд. Сила тока меняется с расстоянием от возбужденного сегмента, декремента.
Механизм распространения возбуждения
Возбуждение охватывает последовательно все отделы нервного волокна. R наруж влияет на скорость распространения возбуждения. В экспериментрах Ходжкина изменили внеклеточную среду на масло, которое имеет большее сопротивление, объем снизился на 30-50%. Эксперимент: нерв помещается на параллельные пластинки из серебра, замыкают с помощью ртутной ванночки, объем проведеним растет на 16-30%. Была подтверждена теория местных токов для безмякотных волокон. В мякотных нерных волокнах механизм проведения другой. Миелин имеет рост сопротивления и снижение емкости, миелиновая оболочка прерывается перехватами Ранвье – сальтаторное проведение. R на 1 см2 поверхности в перехвате Ранвье = 10-20 Ом, в миелиновой оболочке = 0,003-0,005 Ом. Петли тока в миелиновых нервных волокнах выходят через невозбужденый перехват Ранвье, находящийся спереди от возбужденного . Эксперименты Тасаки.
1. Электроды стоят на миелине, два выходящих тока (это токи, выходящие из
последующего и предыдущего перехвата Ранвье. Входящий ток не регистрируется.
2. Средний электрод на перехвате. Появляется входящий ток.
По немиелинизированному волокну ПД распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший ПД за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые ПД. Сами ПД не перемещаются, они исчезают там же, где возникают. Главную роль в возникновении нового ПД играет предыдущий.
По миелинизированному волокну ПД распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье ПД возбуждает 2-ой, 3-ий, 4-ый и даже 5-ый, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Это увеличивает скорость распространения ПД по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na и K , расходующихся в результате возникновения ПД, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.
Электрокинетический потенциал: возникновение, измерение и факторы, определяющие его величину. Примеры электрокинетических явлений, их характеристика и научно-практическое значение.
Потенцил Действия
1. Объясняется поворотом диполя на 1800.
2. Теория альтерации Германа. При возбуждении возникает избыток кислых продуктов, которые несут отрицательны заряд, что приводит к разности потенциалов между возбужденным и невозбужденым учаском.
3. Мембранная теория Берштейна. В возбужденном участке мембраны резко увеличивается проницаемость для всех ионов, концентрации ионов смешиваются и участок становится электронейтральным.
4. Ходжкин и Хаксли. Рост проницаемости мембраны для ионов в месте воздействия. При возбуждении электропроницаемость мембраны увеличивается примерно в 500 раз. Max увеличивается проницаемость мембраны для Na (отсюда Na-теория ПД). Na свободно проходит внуть клетки. При возбужедении электро-химическое равновесие определяется потенциалом Na . Равновесный потенциал для К = –97 мВ, для Na = 50 мВ. При возбужедении мембрана перезаряжается. Положение обратной активации и инактивации Na -каналов, Na -канал может активироваться (открываться) при определенных значениях потенциала. Причина активации Na -каналов – деполяризация мембраны, чем больше деполяризация, тем больше проницаемость мембраны для Na . Зависимость близка к линейной в подкор уровне; как только мембрана достигнет критического уровня деполяризации – зависимость нелинейная, лавинообразный вход Na в клетку.
Электрокинетический потенциал, x-потенциал, дзета-потенциал, часть общего скачка потенциала на границе двух фаз, определяющая относительное перемещение этих фаз при электрокинетических явлениях. Общий скачок потенциала при пересечении межфазной границы в дисперсных системах обусловлен существованием двойного электрического слоя. Электрокинетический потенциал — перепад потенциала по той части диффузного слоя, в пределах которой жидкость может быть вовлечена в тангенциальное движение относительно межфазной поверхности при внешнем воздействии на систему. Под влиянием сильно адсорбирующихся на поверхности ионов или изменения pH жидкости может произойти перемена знака на противоположный («перезарядка» поверхности). Электрокинетический потенциал в изоэлектрической точке равен нулю. он является одной из важнейших характеристик жидких коллоидных систем. Его значение и характер изменения при варьировании параметров электролита, адсорбции на пов-сти разл. в-в и т. п. позволяет судить о структуре граничных слоев, особенностях взаимод. компонентов р-ра с пов-стью, заряде пов-сти и т.д.
1). Для объяснения реполяризации используется положение об инактивации Na -каналов. При приближении потенциала мембраны к равновесному для Na , Na -каналы инактивируются и посупление Na в клетку прекращается. К графику: в основе регенеративный процесс (сам себя поддерживающий), развивающийся по принципу обратной связи.
2). Рост К проницаемости мембраны. Не столь значителен, как для Na ( в 5-15 и 500 раз соответственно). Проницаемость для К развивается медленнее, чем для Na . Ионы К в этой ситуации будут выходить наружу и выносить заряд.
3). Механизм активного транспорта, представленный K-Na-насосом. 3 Na внутрь и 2 К наружу.
Эксперименты Ходжкина и Хаксли.
Гигантский аксон кальмара. Из внеклеточной среды были удалены 2/3 Na . При этом амплитуда ПД снизилась ? на 50%. Замена внутриклеточного Na на другие ионы приводит к некоторому росту ПД. Замена ? внутриклеточного К на Na приводит к значительному снижению ПД.
Метод фиксации потенциала
метод Петч-Клемпинга. С его помощью можно зафиксировать на длительное время значение мембранного потенциала на любом желаемом уровне. Это делается с помощью внешнего генератора напряжения
Суммарные мембранные токи при ПД
1. Подпороговая область:
Слабое изменение мембранного потенциала, суммарный ионный ток направлен от клетки наружу, так как поток К , выходящий из клетки, уже усиливается из-за удаления мембранного потенциала от равновесного потенциала для К . Входящий ток Na еще слаб, так как рост Na -проницаемости пока невелик. Однако с развитием деполяризации Na-ый поток постепенно нарастает.
2. Критический уровень деполяризации:
В этот момент суммарный ионный ток через мембрану равен нулю, так как встречные токи ионов Na и К уравновешивают друг друга. Даже небольшая дальнейшая деполяризация приводит к росту входа Na -тока в сотни раз.
3. Во время фазы деполяризации резко увеличивается Na -проницаемость и суммарный мембранный ток, направленный внутрь клетки. Выходящий К -ток растет медленнее и становится заметным только к моменту пика потенциала.
4. Фаза реполяризации:
В момент пика потенциала большинство Na -каналов инактивированны, а К -ток max. Поэтому суммарный мембранный ток – выходящий.
Кальциевая теория активации и инактивации Na -каналов
В состоянии покоя у наружного отверстия Na -канала находится Са2 , который электростатически тормозит проникновение Na в канал. При возбуждении наружная поверхность мембраны заряжена отрицательно, при этом Са2 уходят со своих мест, вход открывается и Na входит в клетки.
Инактивация: по ходу деполяризации узкие Na -каналы могут закупориваться Na . Во многих каналах есть воротные белки (могут менять свое местоположение под влиянием изменения потенциала). В состоянии покоя активационный белок закрыт, а инактивационный открыт. При возбуждении открывается активационный белок в момент закрывания инактивационного белка. В конце реполяризации белки так же закрываются и потом открываются (исходное состояние).
Факторы, определяющие скорость распространения возбуждения по нервному волокну
1. Пространственная константа определяет величину декремент
деполяризации, ? – пространственная константа.
2. Коэффициент надежности, соотношение между амплитудой ПД
критической энергией и ПП. S=ПД/(Екр–ПП), ПД=120мВ, ПП=–70мВ, Екр=–55мВ ? S=8. Чем больше S, тем быстрее проведение.
3. Временная константа ? мембр. При возбуждении мембраны меняется
заряд. Длительность перезарядки мембраны. ?мембр=Rm*Cm. Чем больше ? мембр, тем ниже С. Vраспр=S*?/?мембр.
§
В основе сокращения мышц лежит взаимное перемещение двух систем нитей, образованных актином и миозином. АТФ гидролизуется в активном центре, расположенном в головках миозина. Гидролиз сопровождается изменением ориентации головок миозина и перемещением нитей актина. Регуляция сокращения обеспечивается специальными Са-связывающими белками, расположенными на нитях актина или миозина. В основе сокращения мышц практически всех типов лежит перемещение двух систем белковых нитей (филаментов), построенных в основном из актина и миозина.
Для высокоэффективного преобразования энергии АТФ в механическую работу мышцы должны обладать строго упорядоченной структурой.
Строение и свойства актина. Мономеры актина (их часто называют G-актином, то есть глобулярным актином) могут взаимодействовать друг с другом, образуя так называемый фибриллярный (или F-актин). Процесс полимеризации можно инициировать повысив концентрацию одно- или двухвалентных катионов или добавив специальные белки. Процесс полимеризации становится возможным потому, что мономеры актина могут узнавать друг друга и образовывать межмолекулярные контакты. Полимеризованный актин внешне похож на две скрученные друг относительно друга нитки бус, где каждая бусина представляет собой мономер актина. Процесс полимеризации актина строго упорядочен, и мономеры актина упаковываются в полимер только в определенной ориентации. Тот конец полимера, где скорость полимеризации больше, называют плюс-концом, а противоположный конец полимера обозначают как минус-конец.
Актин является уникальным строительным материалом, широко используемым клеткой для построения различных элементов цитоскелета и сократительного аппарата. Есть белки, связывающиеся с мономерным актином (например, профилин). Эти белки, находясь в комплексе с глобулярным актином, препятствуют его полимеризации. Есть специальные белки, которые, как ножницы, разрезают уже сформировавшиеся нити актина на более короткие фрагменты. Некоторые белки преимущественно связываются и формируют шапочку (“кепируют”) на плюс-конце полимерного актина. Другие белки кепируют минус-конец актина. Существуют белки, которые могут сшивать уже сформировавшиеся нити актина.
Все нити актина в саркомере имеют постоянную длину и правильную ориентацию, при этом плюс-концы филаментов располагаются в Z-диске, а минус-концы – в центральной части саркомера. Вследствие такой упаковки нити актина, расположенные в левой и правой частях саркомера, имеют противоположную направленность.
Строение и свойства миозина. В состав молекулы миозина скелетных мышц входят шесть полипептидных цепей – две так называемые тяжелые цепи миозина и четыре легкие цепи миозина (ЛЦМ). Эти цепи прочно ассоциированы друг с другом (нековалентными связями) и образуют единый ансамбль, который собственно и является молекулой миозина.
Тяжелые цепи миозина имеют большую молекулярную массу и сильно асимметричную структуру. У каждой тяжелой цепи есть длинный спирализованный хвост и маленькая компактная грушевидная головка. Спирализованные хвосты тяжелых цепей миозина скручены между собой наподобие каната. Этот канат обладает довольно высокой жесткостью, и поэтому хвост молекулы миозина образуют палочкообразные структуры. В нескольких местах жесткая структура хвоста нарушена. В этих местах располагаются так называемые шарнирные участки, обеспечивающие подвижность отдельных частей молекулы миозина. Шарнирные участки легко подвергаются расщеплению под действием протеолитических (гидролитических) ферментов, что приводит к образованию фрагментов, сохраняющих определенные свойства неповрежденной молекулы миозина.
В области шейки, то есть при переходе грушевидной головки тяжелой цепи миозина в спиральный хвост, располагаются короткие легкие цепи миозина. С каждой головкой тяжелой цепи миозина связаны одна регуляторная и одна существенная легкая цепь миозина. Обе легкие цепи миозина тем или иным способом влияют на способность миозина взаимодействовать с актином и участвуют в регуляции мышечного сокращения.
Палочкообразные хвосты могут слипаться друг с другом за счет электростатических взаимодействий. При этом молекулы миозина могут располагаться либо параллельно, либо антипараллельно друг относительно друга.
Хвосты миозина скелетных мышц могут упаковываться как в параллельном, так и в антипараллельном направлении. Комбинация параллельной и антипараллельной упаковок приводит к формированию так называемых биполярных (то есть двухполюсных) филаментов миозина. Половина молекул миозина повернута своими головами в одну сторону, а вторая половина – в другую сторону. Биполярный миозиновый филамент располагается в центральной части саркомера.
Главной “моторной” частью миозина скелетных мышц является головка тяжелой цепи миозина вместе со связанной с ней легкими цепями миозина. Головки миозина могут дотягиваться до нитей актина и контактировать с ними. При замыкании таких контактов образуются так называемые поперечные мостики, которые собственно генерируют тянущее усилие и обеспечивают скольжение нитей актина относительно миозина.
Рассмотрим цикл гидролиза АТФ и перемещение головки по актину. В исходном состоянии головка миозина не насыщена АТФ, актинсвязывающие центры сближены и головка прочно взаимодействует с актином. При связывании АТФ в активном центре “пасть” раскрывается, актинсвязывающие участки удаляются друг от друга, прочность связи миозина с актином ослабевает и головка диссоциирует от нити актина. Гидролиз АТФ в активном центре диссоциировавшей от актина головки миозина приводит к закрыванию щели активного центра. После гидролиза АТФ до АДФ и неорганического фосфата шейка занимает положение, перпендикулярное длинной оси нити актина. После всех этих событий головка миозина вновь оказывается способной взаимодействовать с актином. Образование комплекса с актином вызывает структурные изменения в головке миозина. Эти изменения позволяют выбросить из активного центра миозина неорганический фосфат, который образовался в ходе гидролиза АТФ. Головка миозина проталкивает нить актина на шаг вперед. После этого из активного центра выбрасывается другой продукт реакции, АДФ. Цикл замыкается, и головка переходит в исходное состояние.
Механизмы регуляции мышечного сокращения. Ионы Са2 являются сигналом для начала мышечного сокращения.
Миозиновый тип регуляции сократительной активности. Простейший способ миозиновой регуляции описан для некоторых мышц моллюсков. Миозин моллюсков по своему составу не отличается от миозина скелетных мышц позвоночных. Считается, что при отсутствии Са2 легкие цепи обернуты вокруг шарнирного участка тяжелой цепи миозина. При этом подвижность шарнира сильно ограничена. Головка миозина не может совершать колебательных движений, она как бы заморожена в одном положении относительно ствола толстого филамента. Очевидно, что в таком состоянии головка не может осуществлять колебательные движения и вследствие этого не может перемещать нить актина. При связывании Са2 происходят изменения структуры легких и тяжелых цепей миозина. Резко повышается подвижность в области шарнира. Теперь после гидролиза АТФ головка миозина может осуществлять колебательные движения и проталкивать нити актина относительно миозина.
Оказалось, что с миозиновыми филаментами гладких мышц связан специальный фермент. Этот фермент получил название “киназа легких цепей миозина” (КЛЦМ). В состоянии покоя при низкой концентрации Са2 в цитоплазме киназа легких цепей миозина неактивна. Это связано с тем, что в структуре фермента есть специальный ингибиторный (блокирующий активность) участок. Ингибиторный участок попадает в активный центр фермента и, не давая возможности взаимодействовать с истинным субстратом, полностью блокирует активность фермента.
В цитоплазме гладких мышц есть специальный белок кальмодулин, содержащий в своей структуре четыре Са-связывающих центра. Связывание Са2 вызывает изменения в структуре кальмодулина. Насыщенный Са2 кальмодулин оказывается способным взаимодействовать с КЛЦМ. Посадка кальмодулина приводит к удалению ингибиторного участка из активного центра, и киназа легких цепей миозина как бы просыпается. Только после фосфорилирования легкой цепи миозин оказывается способным взаимодействовать с актином и начинается мышечное сокращение.
Актиновый механизм регуляции мышечного сокращения. Связанный с актином механизм регуляции сократительной активности характерен для поперечнополосатых скелетных мышц позвоночных и сердечной мышцы. Нити фибриллярного актина в скелетных и сердечных мышцах имеют вид двойной нитки бус. Нитки бус актина перекручены друг относительно друга, поэтому с двух сторон филамента образуются канавки. В глубине этих канавок размещается сильно спирализованный белок тропомиозин. Каждая молекула тропомиозина состоит из двух одинаковых (или очень похожих друг на друга) полипептидных цепей, которые перекручены. Располагаясь внутри канавки актина, палочкообразная молекула тропомиозина контактирует с семью мономерами актина.
На актиновом филаменте помимо тропомиозина располагается еще и тропониновый комплекс. В состоянии расслабления концентрация Са2 в цитоплазме очень мала. Регуляторные центры тропонина С не насыщены Са2 . Ингибиторный и С-концевой участки тропонина I взаимодействуют с актином и с помощью тропонина Т выталкивают тропомиозин из канавки на поверхность актина. До тех пор пока тропомиозин располагается на периферии канавки, доступность актина для головок миозина ограниченна. Контакт актина с миозином возможен, но площадь этого контакта мала, вследствие чего головка миозина не может переместиться по поверхности актина и не может генерировать тянущее усилие.
При повышении концентрации Са2 в цитоплазме происходит насыщение регуляторных центров тропонина С. Тропонин С образует прочный комплекс с тропонином I. При этом ингибиторная и С-концевая части тропонина I диссоциируют от актина. Теперь ничто не удерживает тропомиозин на поверхности актина, и он закатывается на дно канавки. Такое перемещение тропомиозина увеличивает доступность актина для головок миозина, увеличивается площадь контакта актина с миозином, и головки миозина приобретают возможность не только контактировать с актином, но и прокатываться по его поверхности, генерируя при этом тянущее усилие.





